An initiative by the Association of Malawians in Ireland (AMAI) has resulted in the securing of €2,500 (about K4,715,114) for the construction of a new house for a needy widow in Dowa whose current hut is almost uninhabitable.
The widow, Florence Tsekani, lives with her seven children and, aside from a decent house, theirs is a day-to-day grim struggle for survival.
AMAI executive chairperson, Henry Mkumbira Phiri, said it was exciting that members have been able to contribute towards the cause within about six days.
“We invited 25 people to contribute €100 each towards the Florence Tsekani house project in Dowa. They have been very willing to support and we have raised the funds within six days,” said Phiri.
He said the construction of the house will begin on July 1, 2024 and end on August 15 2024.
“The money we have raised will cater for a two bedroomed house, a separate kitchen, a toilet and a bathroom. We thought it wise to do the construction after the rainy season ends,” he said.
According to Mkumbira-Phiri, despite living in the diaspora, Malawians in Ireland remain connected to situations at home.
“Malawians in Ireland always remember where they are coming from and it is in our spirit to respond to all issues negatively or positively affecting our families back home,” he said.
Meanwhile, the Dowa district social welfare office which raised an SOS on Tsekani’s house predicament has expressed its vote of thanks to Malawians living in Ireland.
“It is touching how our friends in the diaspora have reacted to this situation. The construction of a new home for Tsekani and her seven children will go a long way in changing their lives,” said the social welfare officer.
Tuberculosis (TB) is a pervasive bacterial infection caused by Mycobacterium tuberculosis (Mtb), contributing significantly to global mortality. Despite Mtb’s latent infection in a substantial proportion of the global population, only a fraction progresses to active TB. The disproportionate burden of TB in Sub-Saharan Africa and Southeast Asia underscores its public health significance.1,2Mycobacterium tuberculosis’s intracellular nature and host-specific interactions highlight the genetic basis of susceptibility.
Numerous studies have explored the connection between genomic loci and TB susceptibility, encompassing candidate genes, family aggregation, twin studies, house contacts, and genome-wide analyses.3 Key players in TB susceptibility include genes encoding proteins such as major histocompatibility complex/human leukocyte antigen (MHC/HLA), tumor necrosis factors (TNFs) and their receptors, immune-related GTPases (IRGs), NRAMP1 (SLC11A1), Toll-like receptors (TLRs), vitamin D nuclear receptor (VDR), and cell surface proteins like collectins.4,5
The complex interplay between host genetics and TB susceptibility underscores the need for a nuanced understanding. While existing literature offers insights, inconsistencies arise, necessitating a deeper exploration of innate immune genes. Genetic polymorphisms associated with TB susceptibility display varying frequencies across populations, suggesting the role of environmental selection pressures.5,6
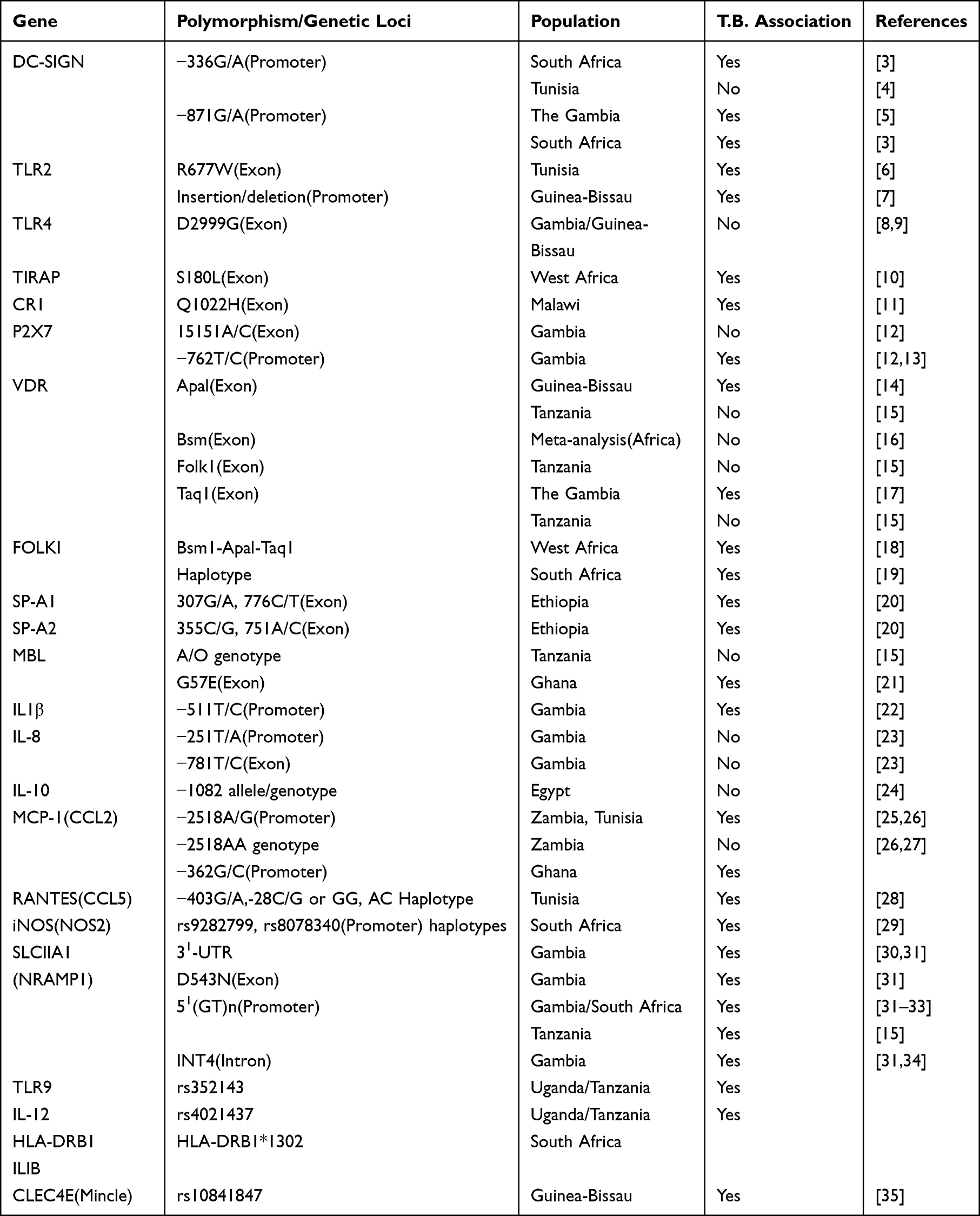
This review focuses on polymorphisms in immune genes linked to pulmonary TB susceptibility in African populations, Table 1. By elucidating the interaction between host molecules and Mtb, we aim to contribute to the design of effective strategies against tuberculosis.
Table 1 Association Studies of Polymorphisms in Immune Genes and Tuberculosis
The African Context
Understanding the genetic basis of tuberculosis (TB) susceptibility is a complex endeavor that requires careful consideration of diverse populations. Focusing on African populations in TB polymorphism studies is a strategic choice driven by various scientific, epidemiological, and ethical considerations. This justification aims to elucidate the rationale behind concentrating exclusively on African populations.
High TB Burden in Africa
Africa bears a substantial burden of TB, with high prevalence rates and a significant impact on public health.7 By concentrating on African populations, we prioritize a region where TB poses a severe threat to community well-being. Tuberculosis in Africa is characterized by a multifaceted interplay of factors, including TB-HIV co-infection, poverty, weak healthcare systems, and drug resistance. These unique challenges underscore the necessity for tailored interventions that address the specific needs of African communities.
Genetic Diversity in African Populations
Africa is characterized by unparalleled genetic diversity, owing to its rich history, diverse ethnicities, and unique population structures.8 Investigating TB polymorphisms in this context allows for a nuanced exploration of genetic variants that may be specific to African populations. This approach aligns with the growing recognition of the importance of population-specific genetic studies.
Prevalence of HIV Co-Infection
TB-HIV co-infection represents a critical aspect of TB epidemiology in Africa. HIV weakens the immune system, heightening susceptibility to TB infection and increasing the risk of TB reactivation among individuals with latent TB. Numerous studies have underscored the strong association between HIV and TB in Africa, with a significant proportion of TB cases occurring among HIV-positive individuals.1,9
Impact of Poverty and Socioeconomic Factors
Poverty and socioeconomic disparities significantly fuel the TB epidemic in Africa. Factors such as poor living conditions, overcrowding, limited healthcare access, and inadequate nutrition contribute to the transmission and persistence of TB in many African communities.10 These challenges exacerbate efforts to control TB and impede the achievement of optimal treatment outcomes.
Challenges Posed by Weak Healthcare Systems
Africa’s healthcare systems grapple with resource constraints, infrastructure deficiencies, and staffing shortages, presenting substantial barriers to effective TB control. Inadequate diagnostic capacities, limited access to quality medications, and suboptimal adherence to treatment regimens further complicate TB management in numerous African countries.11 Strengthening healthcare systems and enhancing access to TB care are paramount for reducing the burden of TB in Africa.
Emergence of Drug Resistance
The emergence of drug-resistant TB strains, including multidrug-resistant (MDR) and extensively drug-resistant (XDR) variants, poses a formidable challenge to TB control efforts across Africa. Limited access to quality diagnostic tools, inappropriate treatment regimens, and poor treatment adherence contribute to the proliferation of drug-resistant TB strains in the region.12 Addressing the menace of drug-resistant TB demands enhanced surveillance, rigorous infection control measures, and expanded access to effective treatment modalities.
Environmental and Socioeconomic Factors
The interplay between genetics and environmental factors is a crucial aspect of TB susceptibility. African populations often face unique environmental challenges and socioeconomic conditions that influence disease dynamics. Investigating TB polymorphisms in this context allows for a holistic understanding of the gene-environment interactions contributing to TB susceptibility.
Precision Medicine and Targeted Interventions
African populations deserve tailored medical interventions that consider their unique genetic makeup. Precision medicine initiatives aim to develop therapies specific to individual genetic profiles.13 By focusing on TB polymorphisms in African populations, we contribute to the development of targeted interventions that address the specific needs of this population. Therefore, the decision to study TB polymorphisms in African populations is rooted in the region’s high TB burden, genetic diversity, unique environmental challenges, and the pursuit of precision medicine. By concentrating our efforts in this context, we aim to unravel population-specific genetic factors influencing TB susceptibility, ultimately contributing to more effective and targeted strategies for TB control in Africa.
Epidemiological Data
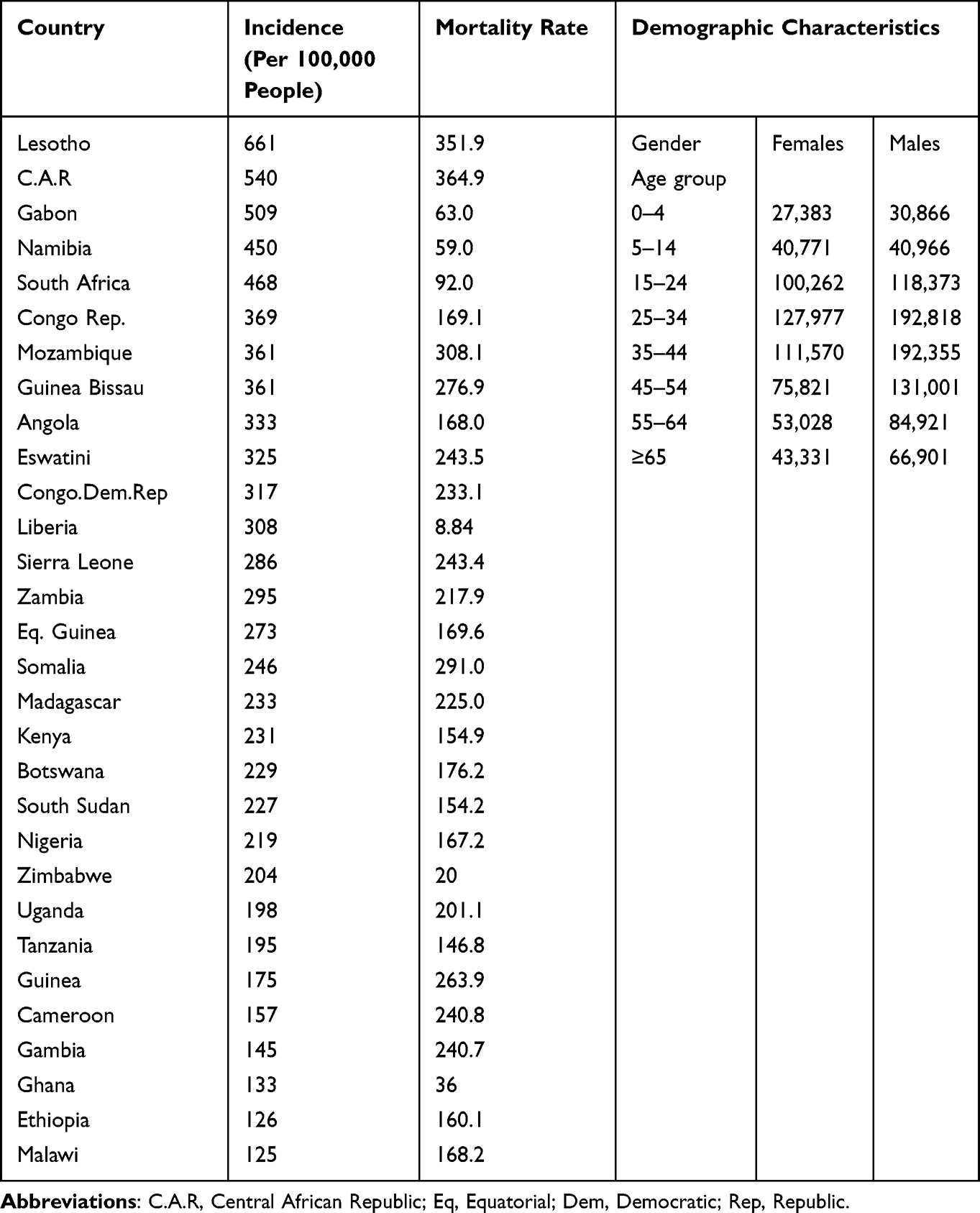
Tuberculosis remains a pressing public health concern, particularly in African populations, where it continues to exert a significant burden on health systems and communities. The incidence rates, mortality rates, and demographic characteristics associated with TB in Africa are shown in Table 2.14
Table 2 The Incidence, Mortality Rates, and Demographic Characteristics Associated with TB in Africa
TB Incidence Rates
Countries such as Lesotho, C.A.R, Gabon, South Africa have the highest Tb incident rates on the continent. However, progress has been made in reducing TB incidence in several African countries through increased access to diagnostic and treatment services, Table 2.
TB Mortality Rates
Somalia, Mozambique, Lesotho and C.A.R are among the countries with the highest TB mortality rates in Africa. TB mortality rates in Africa are closely linked to factors such as HIV co-infection, drug resistance, and healthcare access. While mortality rates have declined in recent years due to improved TB control efforts, they remain significant, particularly in countries with high HIV prevalence.
Demographic Features
Young adults, particularly those in the economically productive age group, are disproportionately affected by TB. Additionally, men are more likely to develop TB than women in many African countries, due to social and behavioral factors as seen in Table 2. Socioeconomic disparities, including poverty, overcrowding, and limited access to healthcare, contribute to the unequal distribution of TB within communities. Urban areas tend to have higher TB burdens due to population density and environmental factors, although rural areas are not exempt from the disease.
Roles of Proteins in Host – Pathogen Interaction
In the realm of tuberculosis susceptibility, extensive research is focused on elucidating the intricate roles of various proteins to decipher the underlying mechanisms governing host-pathogen interaction. These proteins, encompassing Toll-like receptors (TLRs), Dendritic Cell-Specific Intercellular Adhesion Molecule-3 Grabbing Non-Integrin (DC-SIGN), the vitamin D nuclear receptor, soluble C-type lectins such as surfactant proteins A and D, C-type Lectin Domain Family 4 Member E (CLEC4E), and mannose-binding lectin (MBL), represent integral components of the immune system’s response to Mycobacterium tuberculosis (MTB) infection.
As we embark on an in-depth exploration of the nuanced interplay between these proteins and MTB, we intent to uncover their individual and collective contributions to TB susceptibility. Farthermore, discerning the roles of phagocyte cytokines, including Interleukin-1 (IL-1), Interleukin-6 (IL-6), Interleukin-10 (IL-10), Interleukin-12 (IL-12), and Interleukin-18 (IL-18), alongside chemokines such as Interleukin-8 (IL-8), monocyte chemoattractant protein 1 (MCP-1), and Regulated upon activation, normal T-cell expressed and secreted (RANTES). This comprehensive investigation serves to enhance our understanding of the immune response to TB infection, paving the way for novel insights into TB susceptibility mechanisms and potential avenues for therapeutic intervention.
The Human Leukocyte Antigens(HLAs)
The Human Leukocyte Antigen (HLA) molecules are proteins found on the surface of cells in the human body. They play a crucial role in the immune system by presenting antigens, which are fragments of proteins from pathogens like bacteria and viruses, to T cells. This process is essential for the immune system to recognize and respond to foreign invaders while distinguishing them from the body’s own cells. HLA molecules are highly diverse and polymorphic, meaning they exist in many different forms within human populations. This diversity is important for immune system function and contributes to individual differences in immune responses to pathogens, susceptibility to diseases, and compatibility for organ and tissue transplantation.
Recent studies have revealed that specific Human Leukocyte Antigen (HLA) alleles might contribute to either increased or decreased susceptibility to tuberculosis (TB) within African communities.5 For instance, investigations have established correlations between certain HLA class I alleles—such as HLA-B39:01 and HLA-B58:01—and elevated TB susceptibility among West African populations. Conversely, alternative HLA class I alleles, like HLA-B*57:01, have demonstrated a protective association against TB within African cohorts.15,16
Furthermore, HLA class II alleles have emerged as significant factors in TB susceptibility among African populations.16 Notably, research has identified links between particular HLA-DRB1 alleles—such as HLA-DRB115:01 and HLA-DRB104:05—and heightened susceptibility to TB among East and West African groups. Conversely, contrasting HLA-DRB1 alleles, including HLA-DRB110:01 and HLA-DRB111:01, have shown protective effects against TB within African populations.
It is crucial to recognize the intricate nature of the relationship between HLAs and TB susceptibility, influenced by diverse factors such as genetic background, environmental exposures, and host-pathogen interactions. Moreover, the prevalence of specific HLA alleles may exhibit variability across different African populations, underscoring the necessity for population-specific investigations to comprehensively grasp the genetic determinants of TB susceptibility.
Soluble C-Type Lectins (Collectins)
Collectins, including MBL, SP-A, and SP-D, are calcium-dependent lectins crucial for innate immunity. Previous data on calcium-dependent (C type) lectins reveals the close relationship between the bacillus with MBL and SP-A, SP-D17 Specifically, SP-A and SP-D, located in the alveoli and respiratory epithelium, play a pivotal role in the defense against intracellular pathogens, including Mtb. SP-A and SP-D, collagen-like glycoproteins, serve as opsonins, enhancing phagocytosis by alveolar macrophages.17–19 Additionally, SP-A interacts with alveolar macrophages, modulating TLR2 and TLR4 pathways,18 showcasing the intricate immune pathways influenced by these collectins. SP-A’s impact on inflammatory cells depends on the cell state, resulting in the inflammatory paradox. While enhancing cytokine release during infection, SP-A also suppresses macrophage secretion, revealing a complex regulatory role in the immune response.
Polymorphic changes in the genes encoding SP-A and SP-D, particularly in the amino acid sequences, contribute to susceptibility to TB.20 Genetic changes in SP-A and SP-D amino acid sequences have functional consequences. Single nucleotide polymorphisms in these proteins occur when a single nucleotide (adenine, thymine, cytosine, or guanine) in a DNA sequence is altered or substituted with a different single nucleotide at a specific position in the genome. These genetic variants may either result in a synonymous or non-synonymous substitution. Since interactions of SP-A and SP-D with the same and or different pathogens occur. A polymorphism in SP-A reduces binding to Mtb, increasing susceptibility, while a genetic change in SP-D alters the sequence, affecting bacterial surface binding and increasing uptake by alveolar macrophages.
Membrane Binding Lectins (MBL)
Membrane-binding lectin, a soluble pathogen recognition receptor, plays a pivotal role in host defense against bacterial pathogens. Its hexameric structure, akin to the C1q component of the complement system, facilitates the opsonization of microbes, enhancing phagocytosis. MBL also contributes to inflammation by clearing apoptotic cells and releasing cytokines.21,22 MBL gene polymorphisms contribute to variations in blood levels and hexamer formation. MBL gene duplication results in the identification of two genes, MBL1 and MBL2, with MBL2 being the representative in humans. The MBL2 gene encompasses four coding regions (exons) separated by three non-coding regions (introns) at codon positions 52, 54, and 57, featuring variant alleles, D, B, and C, respectively. Certain alleles are linked to impaired hexamer formation and reduced blood levels.5
Mutations at specific codons resulting in low MBL levels increase susceptibility to various infections, particularly in individuals with other immune defects. Despite numerous studies exploring MBL alleles and their polymorphisms (Table 1), inconsistent results underscore the imperative to comprehensively understand the relationship between TB establishment and the polymorphic variants in the MBL gene.
Mannose-binding lectin-2 variants exhibit relatively frequent occurrences, albeit unevenly distributed across human populations.23 Notably, allele B is prevalent in Asian and native South African populations but absent in the West African population. Conversely, allele D is confined to North Africa and Caucasian populations, while allele C is predominant in Sub-Saharan Africa. These allele frequency disparities shed light on the biological advantage of MBL low levels in specific populations.
In summary, individuals with low MBL levels face increased susceptibility to various infections, particularly when coupled with other immune defects. Despite numerous studies exploring MBL alleles and their polymorphisms (Table 1), inconsistent results underscore the imperative to comprehensively understand the relationship between TB establishment and the polymorphic variants in the MBL gene. Notably, the allele frequency disparities observed, especially the prevalence of allele B in certain populations and the restriction of allele D to specific regions, shed light on the biological advantages associated with MBL low levels in distinct populations. Further investigations are warranted to unravel the precise mechanisms linking MBL polymorphisms and TB establishment, paving the way for targeted interventions and a deeper understanding of host-pathogen interactions.
CLEC4E (C-Type Lectin Domain Family 4 Member E)
CLEC4E is a calcium-dependent lectin characterized by carbohydrate recognition domains (CRDs), facilitating its involvement in pathogen recognition and cellular interactions.24 Specifically, CLEC4E encodes the receptor Mincle, situated on the outer membrane of macrophages. Mincle is responsible for recognizing trehalose-6, 6-dimycolate (TDM), a component in the mycobacterial cell wall.25 This recognition triggers signaling cascades within the innate immune system, initiating a robust response to mycobacterial infection.
A single nucleotide polymorphism (SNP) at rs10841847 within CLEC4E has emerged as a crucial determinant in TB susceptibility. The minor allele (G˃A) at this locus is associated with an increased vulnerability to pulmonary TB. This emphasizes the significance of CLEC4E in the context of TB pathogenesis.25 However, the complex landscape of TB susceptibility is likely influenced by multiple genetic factors, necessitating further investigations. While the association between CLEC4E and pulmonary TB is evident, published studies present inconsistent findings. Notably, research in admixed South African populations did not yield statistically significant results, while a study in Northern China reported a decreased predisposition to active TB associated with CLEC4E minor alleles.24,26,27 These discrepancies underscore the need for more comprehensive research to establish the causal link between CLEC4E and TB susceptibility, considering potential confounding factors.
Studies involving mice lacking the Mincle gene offer insights into its role in the immune response against Mycobacterium tuberculosis (Mtb). Depletion of Mincle resulted in the inability to recruit and adhere neutrophils to the site of invasion, impacting the formation of lung granulomas.28 Altered DNA sequences, such as those associated with the rs10841847 SNP, disrupt the normal functioning of Mincle receptors, potentially compromising the ability to bind tubercle glycolipid and increasing susceptibility to TB. Interestingly, studies with knockout mice showed a paradoxical outcome, with a higher bacterial load compared to wild-type mice, suggesting a Mincle-mediated immune response against Mtb.29 This paradox may be attributed to additional genetic factors influencing the sustained immune response, indicating the complexity of host-pathogen interactions in secondary TB.
Pathogen Recognition Receptors (PRRs)
Pathogen recognition receptors are membrane and transmembrane receptors located on the dendritic cells, macrophages, and epithelial cells (exogenous sensors) that recognize different types of Pathogen Associated Molecular Patterns (PAMPs). Macrophage PRRs and phagocytic receptors causative to mycobacterial diseases, especially T.B., have been discussed in several recent reviews.17,24,25 These include cell membrane-bound receptors, such as the mannose receptor (M.R., CD206), dendritic cell-specific ICAM-3-grabbing non-integrin (DC-SIGN) (CD209), Dectin-1, and TLRs. However, in this review, we shall focus on the DC-SIGN and TLRs, which have been associated with Tuberculosis in the African population. This is because they have been adequately studied and have been associated with tuberculosis disease susceptibility and are expressed on many immune cells that recognize numerous molecules of different pathogens including the Mycobacterium tuberculosis.
The exploration of Pathogen Recognition Receptors (PRRs) in dendritic cells, macrophages, and epithelial cells is crucial for deciphering how these receptors recognize diverse Pathogen Associated Molecular Patterns (PAMPs). The emphasis on cell membrane-bound receptors, such as mannose receptor (M.R., CD206), dendritic cell-specific ICAM-3-grabbing non-integrin (DC-SIGN), and Toll-Like Receptors (TLRs), provides a foundation for understanding the initial steps of host-pathogen recognition.30–32
The selective focus on DC-SIGN and TLRs, particularly TLR2, TLR4, and TLR9, is well-justified, considering their extensive associations with TB susceptibility in the African population.31,33,34 These receptors, expressed on various immune cells, serve as critical mediators in the immune response against Mycobacterium tuberculosis. Their well-studied roles in recognizing M. tuberculosis components make them pivotal players in the host defense against TB.35,36
The investigation of Toll-Like Receptors (TLRs) further deepens our understanding of the genetic variants associated with TB susceptibility. The elucidation of TLR1, TLR2, TLR4, and TLR9 variations across different populations provides a nuanced perspective on the global landscape of TB genetics.31 Polymorphisms in these TLRs, including TLR1 G1805T, TLR2 T597C, T1350C, TLR6 C745T, and TLR4 Asp 299Gly, Thr399Ile, reveal potential risk factors for TB susceptibility.31,33,34,37
The signaling pathways initiated by TLR2 and TLR4, leading to the activation of NF-κB and other cascades, are critical in understanding TB susceptibility.35 The identification of specific genetic changes in TLR2, TLR4, and TLR9 associated with TB susceptibility, such as the R753Q variant in TLR2, adds a layer of complexity to our understanding.38 The review delves into the influence of these variants on signaling pathways and their impact on TB progression.
Toll-Like Receptors (TLRs)
Toll-like receptors expressed on various immune cell types, serve as critical mediators in the immune response against a spectrum of pathogens, including Mtb.31,39 Recognizing a diverse array of microbial molecules, TLRs initiate transcription and proinflammatory gene processes. TLRs are categorized based on their cellular localization—surface-bound (eg, TLR2 and TLR4) and intracellular (eg, TLR8 and TLR9).35 The mycobacterial cell wall components are recognized by multiple TLRs, emphasizing their crucial role in the immune response to MTB.34,40
Genetic variants within TLRs have been identified globally, with polymorphisms in TLR1, TLR2, TLR4, TLR8, and TLR9 associated with TB susceptibility.26,31,40 Notably, TLR2 stands out with higher prevalence in genetic variants, forming a heterodimer with TLR1 or TLR6.36 The TLR2 signaling pathway, particularly involving the TIR domain-containing adaptor protein (TIRAP), plays a pivotal role in the inflammatory response to MTB.35,36
In the context of the African population, TLR-2, TLR-4, and TLR-9 emerge as key players in TB susceptibility. The R753Q variant in TLR2 influences disease progression in children.38 Contrasting reports on TLR4 variant D299G highlight its association with TB susceptibility in HIV-infected patients in Tanzania.37,41 The intricate interplay between HIV and TLR variants underscores the multifactorial nature of susceptibility in co-infected individuals. The signaling pathways initiated by TLR2 and TLR4, culminating in the activation of NF-κB, are central to TB susceptibility. Noteworthy changes in these domains correlate with variations in TB susceptibility. The nonsynonymous SNP S180L in TIRAP confers protective signal transduction within TLR2.10 Additionally, TLR7 and TLR8 genetic polymorphisms are associated with TB susceptibility, influencing phagocytosis and immune activation.42 Evidence from TLR9-deficient mice further supports the critical role of TLRs in the innate defense against MTB.26 These results provide proof that TLRs and their signaling moieties in their pathways possess susceptible genes that are associated with Tuberculosis. However, due to variations within individual genes in the different racial populations, it is very hard to make a close linkage of the genetic factors that bring about the association.
DC-SIGN is characterized by a carbohydrate recognition domain (CRD) at its extracellular COOH-terminal end. Its expression on macrophages is influenced by tissue type and activation state.32,39 Notably, DC-SIGN is induced in alveolar macrophages from M. tuberculosis-infected patients, marking its relevance in the context of TB infection.35 The binding of M. tuberculosis to DC-SIGN initiates phagocytosis, allowing the pathogen to enter human monocyte-derived D.C.s. This interaction directly interferes with dendritic cell maturation, contributing to the intricate host-pathogen dynamics.33 The cytoplasmic tail of DC-SIGN, housing three conserved motifs, plays a crucial role in ligand binding, receptor signaling, phagocytosis, and intracellular trafficking of ligand particles.33 This calcium-dependent lectin, known for its affinity for mannose-rich molecules, is expressed predominantly on dendritic cells (D.C.s) and, to varying degrees, on macrophages.34 The dynamic interplay between TB and DC-SIGN involves ligand binding, phagocytosis, and modulation of dendritic cell maturation.32,39
Genetic variations in DC-SIGN have been implicated in TB susceptibility, with polymorphisms in the promoter region (eg, −871A/G and −336A/G) associated with distinct susceptibility patterns (Table 1). Notably, studies in the untranslated regions (UTRs) of CD209 and CD207R genes have revealed variations in gene frequency within African indigenous individuals and the Caucasian American population, underscoring the importance of ethnic, racial, and geographical diversity in TB susceptibility assessment.36 A genetic comparison between African and Eurasian populations has revealed notable disparities, particularly in the frequency of the polymorphic allele −871G. This allele is significantly more prevalent in Eurasians than in Africans, suggesting a genetic migration and close association leading to its introduction into the South African population.43 The absence of haplotypes −871G and −336A in most African populations, except for colored South Africans, further emphasizes the impact of genetic migration on TB susceptibility. In conclusion, such genetic variations tend to influence translation and gene polymorphisms.
Solute Carrier Family 11A, Member 1, SLC11A1 or Natural Resistance-Associated Macrophage Protein 1, or NRAMP1)
The SLC11A1 (NRAMP1) gene encodes a membrane transporter protein whose function is to regulate the cytoplasmic cation levels, especially iron. Iron is essential for the cell to generate reactive oxygen and nitrogen intermediates. It, therefore, acts as a divalent cation transporter across phagosomal membranes.38 The origin of the gene from a mouse is responsible for its resistance to infections by intracellular pathogens, especially the mycobacteria.37
SLC11A1’s function as a divalent cation transporter across phagosomal membranes is critical in modulating the concentration of metal ions within the phagosome. This regulation is paramount for macrophage defense against bacterial infections, specifically by reducing the metal ion concentration where bacterial pathogens reside. A defective NRAMP gene, however, results in increased metal ion accumulation, supporting bacterial growth and compromising the host’s ability to resist infection.
Studies across diverse populations and recent meta-analyses have highlighted the influence of SLC11A1 polymorphisms on TB susceptibility. Four key polymorphisms (3=-UTR, D543N, 5= (G.T.) and INT4 have been extensively studied globally. Although findings exhibit some contradictions in various populations, a meta-analysis in China and an association in Turkey underscore the significance of these polymorphisms in TB development.
A systematic review conducted in China further demonstrates the role of SLC11A1 polymorphisms, particularly at the D543N and INT4 loci, in response to infection rather than solely to Mycobacterium tuberculosis.42,44,45 Additionally, studies in the Gambia suggest potential links between SLC11A1 gene polymorphisms, abnormal iron levels, gene-nutrient interactions, and mortality prediction in HIV-infected patients. This raises intriguing questions about the potential risk of TB development in HIV patients with clinically abnormal iron levels and gene-nutrient interactions. Genetic interactions, such as those observed between TLR2 and polymorphism rs3731865 in African Americans, emphasize the significance of gene-gene interactions in host-pathogen genetic studies. These findings highlight the complexity of allelic heterogeneity within SLC11A1 and underscore the importance of considering various factors in understanding TB susceptibility.
Highlighted associations with SNPs in SLC11A1 are attributed to adjacent SNPs in linkage disequilibrium with reported variants. However, the 5- (G.T.) repeat in the promoter region of SLC11A1 has been shown to affect promoter activity, with allele 3 displaying greater basal and lipopolysaccharides (LPS)-stimulated activity. This intriguing gene activity, although insufficient for establishing clear evidence for in vivo activity, links high activity with resistance and low activity with susceptibility to tuberculosis.46
Chemokines (Chemotactic Cytokines)
Chemokines are a large family of structurally homologous cytokines, secreted proteins that signal through cell surface G protein-coupled heptahelical chemokine receptors that stimulate leukocyte movement and regulate the migration of leukocytes from the blood to tissues. Therefore, chemokines are proinflammatory cytokines of low molecular size, which coordinate the migration and activation of different leukocyte populations. The structurally homologous chemokines are categorized into four groups based on conserved cysteine arrangement: CXC, CC, C, and CX3C. These low molecular-size proinflammatory cytokines play a pivotal role in coordinating the migration and activation of various leukocyte populations.1
Upon activation of alveolar macrophages by intracellular M. tuberculosis, a cascade of chemokines is released. Notable among these are IL-8 (CXCL8), monocyte chemoattractant protein 1 (MCP-1, CCL2), RANTES (CCL5), and CXCL10 (IP-10). These chemokines act as molecular messengers, orchestrating the migration and activation of T cells and macrophages to sites of Mtb infection. This orchestrated response forms the characteristic tuberculous granuloma, a fortress against the dissemination of the organism.2 Systematic reviews and research articles have extensively documented the significance and genetic variations within various chemokines combating Mycobacterium tuberculosis.25,47,48 Polymorphic variations, particularly in the promoter regions of these genes, can modulate transcription and translation, influencing the overall production and activity of chemokines.3,4
The production of chemokines is indispensable for recruiting inflammatory cells to the infection site. Early recruitment of macrophages is pivotal for infection control.5 M. tuberculosis, a potent inducer of chemokines, triggers protective and immunopathogenic host responses. Studies on human macrophages infected in vitro have revealed the induction of CCL2, CCL3, CCL4, and CCL5 in response to virulent mycobacterial strains.49 Regulated upon activation, normal T-cell expressed, and secreted (RANTES) plays a critical role in T-cell activation and macrophage activation.50 Clinical studies have detected the presence of MCP-1, MIP-1a, RANTES, and IP-10 in the serum and bronchoalveolar lavage of TB patients.51 Alveolar macrophages, in particular, produce a distinct profile of chemokines, suggesting a role in the influx of cells to the infection site, and influencing granuloma formation.52 The expression of chemokines by macrophages not only influences TNF-α production but also affects chemokine receptor expression. Specifically, CCL2, CCL3, CCL4, CCL5, CXCL10, and CXCL13 expression is modulated by macrophages in response to Mtb infection.53 Furthermore, TNF-α itself can influence the chemokine network expression, indirectly impacting granuloma formation.54 This intricate interplay extends to mycobacterial cell wall components, regulating the induction of chemokine secretion by macrophages.55
Rantes (Ccl5)
RANTES (regulated upon activation, normal T-cell expressed, and secreted), a member of the C-C chemokine subfamily or beta chemokine. RANTES exhibits chemotactic properties for monocytes, memory T cells, and eosinophils. In addition, it triggers histamine release from human basophils.56,57 Notably, in a murine model, RANTES has been identified as a key player in promoting granuloma formation within M. tuberculosis-infected lungs.56 This process involves the recruitment of immune cells to the infected area, subsequently limiting the intracellular growth of mycobacteria and inducing direct antimicrobial activity, notably through the release of nitric oxide (NO) in alveolar macrophages.58 The protective immunity conferred by RANTES, as evidenced in knockout mice, underscores its significance in the defense against TB.20
Exploring the genetic landscape, the promoter region of the RANTES gene reveals two variants, −403G/A and −28C/G, associated with Tuberculosis (Table 1). These single nucleotide polymorphisms may confer either resistance or susceptibility to TB.59 A recent discovery highlights a functionally important polymorphism, In1.1T/C, located in the intron of RANTES. This polymorphism regulates gene transcription by differentially binding to alternative forms of nuclear proteins. The complexity of RANTES transcription regulation is further unraveled through haplotype analysis. Haplotypes II (A-C-T) and V (G-C-C) exhibit opposing effects on RANTES transcription, with II up-regulating and V down-regulating the process. The luciferase assay, correlating RANTES SNPs with transcription activity, supports this functional insight. Consequently, RANTES concentrations in bronchoalveolar lavage fluid of pulmonary TB patients demonstrate acute elevation during infection and subsequent decline during convalescence. This underscores the dynamic role of RANTES up-regulation in the immune response against M. tuberculosis infection.
Mcp-1 (Ccl2)
The monocyte chemoattractant protein-1, or MCP-1, is a β chemokine and a potent chemotactic factor for monocytes and macrophages. During lung infection by M. tuberculosis, the human monocytes within the body are stimulated to release MCP-1.60 During the host M. tuberculosis-specific immune response, MCP-1 participates in the inhibition of the in vivo dissemination of M. Tuberculosis regulating the secondary immune response. Therefore, MCP-1 is a vital promoter and regulator of a host’s defense against Tuberculosis infection. However, experimental studies in mice have shown that a deficiency in the chemokine MCP-1 alters the monocyte recruitment and formation of the granuloma, and yet it has the advantage of decreased M. Tuberculosis susceptibility.41 Studies on genotype and allele frequencies especially at the locus −2518 have shown contradictory results and have not yielded concrete and adequate proof for the effects of the polymorphisms in the MCP-1 gene. However, the carriers of the −2518 GG genotype were reported to produce high levels of MCP-1, which inhibits IL-12 production in response to M. Tuberculosis and promotes active pulmonary T.B.,61 whereas Thye et al62 initially found the association with the −2518G allele with T.B. resistance in Ghana, and further genotyping analysis using Microarray technology identified −362C allele as the only true protective variant of MCP-1, with a linkage disequilibrium in the region of −2518 and-362. This study is a deterrent note for the need to carry out a comprehensive genetic investigation to search for the actual cause of the polymorphism that exists in the variant and gene of interest.
Conclusion
In summary, the exploration of genetic factors influencing tuberculosis susceptibility in African populations represents a significant step forward in our understanding of the complex interplay between host genetics and infectious diseases. The comprehensive analysis presented in this article delves into the polymorphisms within immune genes and their association with TB susceptibility specifically within the African context. This study contributes valuable insights into the genetic landscape of TB susceptibility in African populations. By focusing on this region, the research sheds light on population-specific genetic variants that may play a pivotal role in determining an individual’s susceptibility to TB.
One key research gap pertains to the lack of comprehensive studies that consider the diverse genetic, environmental, and socio-economic factors influencing TB outcomes across different African populations. Existing research often fails to adequately account for the genetic diversity present within African populations, leading to inconsistent findings and limited generalizability of results. Moreover, there is a scarcity of longitudinal studies that can elucidate the temporal dynamics of TB susceptibility and progression in African populations, hindering our understanding of the disease’s trajectory and the efficacy of interventions over time.
Additionally, the reliance on small sample sizes and single-center studies limits the statistical power and reliability of findings, further exacerbating the inconsistency of outcomes observed in earlier research efforts. Furthermore, there is a scarcity of studies that integrate genetic, environmental, and socio-economic data to comprehensively assess the determinants of TB susceptibility in African populations, impeding our ability to develop targeted interventions tailored to the specific needs of these populations.
To address these research gaps and enhance our understanding of TB susceptibility in African populations, we propose several suggestions for further similar research. Firstly, there is a need for larger, multicenter studies that encompass diverse African populations to ensure the generalizability of findings and account for population-specific factors. Such studies should employ longitudinal designs to capture the temporal dynamics of TB susceptibility and progression, allowing for the identification of predictive biomarkers and the evaluation of interventions over time.
Moreover, future research efforts should prioritize the integration of genetic, environmental, and socio-economic data through interdisciplinary collaborations between geneticists, epidemiologists, clinicians, and social scientists. This holistic approach will provide a more nuanced understanding of the complex interactions underlying TB susceptibility in African populations and inform the development of targeted interventions that address the multifactorial nature of the disease.
Furthermore, leveraging emerging technologies such as genomic sequencing, big data analytics, and machine learning algorithms can facilitate the identification of novel genetic determinants and biomarkers associated with TB susceptibility in African populations. By harnessing these innovative approaches, researchers can overcome the limitations of earlier studies and advance our understanding of TB epidemiology in Africa, ultimately contributing to the development of effective strategies for TB prevention, diagnosis, and treatment tailored to the specific needs of African populations.
Abbreviations
MHC, Major histocompatibility complex; SLC11A1, Solute carrier family 11a member 1; HLA, Human leukocyte antigen; VDR, Vitamin D nuclear receptor; TNF, Tumour necrosis factor; IRGM, Immunity-related GTPase family M protein; SNP, Single Nucleotide Polymorphism; MTBC, Mycobacterium tuberculosis complex; MCP-1, Monocyte chemoattractant protein 1; MBL2, Mannose-binding lectin 2 gene; Mtb, Mycobacterium tuberculosis; NRAMP1, Natural resistance Associated Macrophage Protein 1 gene; SP-A, Surfactant protein A; SP-D, Surfactant protein D; CRD, Carbohydrate recognition domain; RANTES, Regulated upon activation, normal T-cell expressed, and secreted; TB, Tuberculosis; TLR, Toll-like receptor; DC-SIGN, Dendritic Cell-Specific Intercellular adhesion molecule-3 grabbing non-integrin; WHO, World Health Organization; PRR, Pathogen Recognition Receptor; PAMP, Pathogen Associated Molecular Pattern; CLEC4E, C-type Lectin Domain Family 4 Member E; IL, Interleukin; NO, Nitric Oxide.
Data Sharing Statement
All data generated by this manuscript has been integrated into the manuscript.
Acknowledgments
We are thankful to the Makerere University Library for the provision of electronic information resources that enabled us to complete the review article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The authors declare that this research project was funded by the Africa Centre of Excellence in Materials, Product Development & Nanotechnology; MAPRONANO ACE, Grant No. P151847IDA credit 5797-UG, College of Engineering Design Art and Technology, Makerere University.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Harding E. WHO global progress report on tuberculosis elimination. Lancet Respir Med. 2020;8(1):19. doi:10.1016/S2213-2600(19)30418-7
2. Dye C, Reeder JC, Terry RF. Research for universal health coverage. Sci Transl Med. 2013;5(199):199ed13. doi:10.1126/scitranslmed.3006971
3. Barreiro LB, Neyrolles O, Babb CL, et al. Promoter variation in the DC-SIGN-encoding gene CD209 is associated with tuberculosis. PLoS Med. 2006;3(2):e20. doi:10.1371/journal.pmed.0030020
4. Ben-Ali M, Barreiro LB, Chabbou A, et al. Promoter and neck region length variation of DC-SIGN is not associated with susceptibility to tuberculosis in Tunisian patients. Hum Immunol. 2007;68(11):908–912. doi:10.1016/j.humimm.2007.09.003
5. Yim JJ, Selvaraj P. Genetic susceptibility in tuberculosis. Respirology. 2010;15(2):241–256. doi:10.1111/j.1440-1843.2009.01690.x
6. Pepperell CS, Casto AM, Kitchen A, et al. The role of selection in shaping diversity of natural M. tuberculosis populations. PLoS Pathog. 2013;9(8):e1003543. doi:10.1371/journal.ppat.1003543
7. World Health Organization, Global Tuberculosis Report; 2020.
8. Campbell MC, Tishkoff SA. African genetic diversity: implications for human demographic history, modern human origins, and complex disease mapping. Annu Rev Genomics Hum Genet. 2008;9(1):403–433. doi:10.1146/annurev.genom.9.081307.164258
9. Gray JM, Cohn DL. Tuberculosis and HIV coinfection. Semin Respir Crit Care Med. 2013;34(1):32–43. doi:10.1055/s-0032-1333469
10. Harling G, Ehrlich R, Myer L. The social epidemiology of tuberculosis in South Africa: a multilevel analysis. Soc Sci Med. 2008;66(2):492–505. doi:10.1016/j.socscimed.2007.08.026
11. Oga-Omenka C, Tseja-Akinrin A, Sen P, et al. Factors influencing diagnosis and treatment initiation for multidrug-resistant/rifampicin-resistant tuberculosis in six sub-Saharan African countries: a mixed-methods systematic review. BMJ Glob Health. 2020;5(7):e002280. doi:10.1136/bmjgh-2019-002280
12. Andrews JR, Shah N, Gandhi N, et al. Multidrug-resistant and extensively drug-resistant tuberculosis: implications for the HIV epidemic and antiretroviral therapy rollout in South Africa. J Infect Dis. 2007;196(Suppl s3):S482–90. doi:10.1086/521121
14. Chakaya J, Khan M, Ntoumi F, et al. Global Tuberculosis Report 2020 – Reflections on the Global TB burden, treatment and prevention efforts. Int J Infect Dis. 2021;113(Suppl 1):S7–s12. doi:10.1016/j.ijid.2021.02.107
15. Selvaraj P, Uma H, Reetha AM, et al. HLA antigen profile in pulmonary tuberculosis patients & their spouses. Indian J Med Res. 1998;107:155–158.
16. Singh SP, Mehra NK, Dingley HB, et al. Human leukocyte antigen (HLA)-linked control of susceptibility to pulmonary tuberculosis and association with HLA-DR types. J Infect Dis. 1983;148(4):676–681. doi:10.1093/infdis/148.4.676
17. Torrelles JB, Torrelles J, Azad A, et al. Role of C-type lectins in mycobacterial infections. Curr Drug Targets. 2008;9(2):102–112. doi:10.2174/138945008783502467
18. Beharka AA, Gaynor CD, Kang BK, et al. Pulmonary surfactant protein A up-regulates activity of the mannose receptor, a pattern recognition receptor expressed on human macrophages. J Immunol. 2002;169(7):3565–3573. doi:10.4049/jimmunol.169.7.3565
19. Ferguson JS, Voelker DR, McCormack FX, et al. Surfactant Protein D Binds to Mycobacterium tuberculosis Bacilli and Lipoarabinomannan via Carbohydrate-Lectin Interactions Resulting in Reduced Phagocytosis of the Bacteria by Macrophages1. J Immunol. 1999;163(1):312–321. doi:10.4049/jimmunol.163.1.312
20. Salam N, Gupta S, Sharma S, et al. Protective immunity to Mycobacterium tuberculosis infection by chemokine and cytokine conditioned CFP-10 differentiated dendritic cells. PLoS One. 2008;3(8):e2869. doi:10.1371/journal.pone.0002869
21. Vieira F, Kung JW, Bhatti F. Structure, genetics and function of the pulmonary associated surfactant proteins A and D: the extra-pulmonary role of these C type lectins. Ann Anat. 2017;211:184–201. doi:10.1016/j.aanat.2017.03.002
22. Naqvi KF, Endsley JJ. Myeloid C-type lectin receptors in tuberculosis and HIV immunity: insights into co-infection? Front Cell Infect Microbiol. 2020;10:263. doi:10.3389/fcimb.2020.00263
23. Garred P, Larsen F, Seyfarth J, et al. Mannose-binding lectin and its genetic variants. Genes Immun. 2006;7(2):85–94. doi:10.1038/sj.gene.6364283
24. Jo EK. Mycobacterial interaction with innate receptors: tLRs, C-type lectins, and NLRs. Curr Opin Infect Dis. 2008;21(3):279–286. doi:10.1097/QCO.0b013e3282f88b5d
25. Sasindran SJ, Torrelles JB. Mycobacterium Tuberculosis Infection and Inflammation: what is Beneficial for the Host and for the Bacterium? Front Microbiol. 2011;2:2. doi:10.3389/fmicb.2011.00002
26. Carvalho NB, Oliveira FS, Duraes FV, et al. Toll-like receptor 9 is required for full host resistance to mycobacterium avium infection but plays no role in induction of Th1 Responses. Infect Immun. 2011;79(4):1638–1646. doi:10.1128/IAI.01030-10
27. Bowker N, Salie M, Schurz H, et al. Polymorphisms in the pattern recognition receptor mincle gene (CLEC4E) and association with tuberculosis. Lung. 2016;194(5):763–767. doi:10.1007/s00408-016-9915-y
28. Lai YF, Lin T-M, Wang C-H, et al. Functional polymorphisms of the TLR7 and TLR8 genes contribute to Mycobacterium tuberculosis infection. Tuberculosis. 2016;98:125–131. doi:10.1016/j.tube.2016.03.008
29. Mortaz E, Adcock IM, Tabarsi P, et al. Interaction of Pattern Recognition Receptors with Mycobacterium Tuberculosis. J Clin Immunol. 2015;35(1):1–10. doi:10.1007/s10875-014-0103-7
30. Olvany JM, Sausville LN, White MJ, et al. CLEC4E (Mincle) genetic variation associates with pulmonary tuberculosis in Guinea-Bissau (West Africa). Infect Genet Evol. 2020;85:104560. doi:10.1016/j.meegid.2020.104560
31. Zhang Y, Jiang T, Yang X, et al. Toll-like receptor −1, −2, and −6 polymorphisms and pulmonary tuberculosis susceptibility: a systematic review and meta-analysis. PLoS One. 2013;8(5):e63357. doi:10.1371/journal.pone.0063357
32. Puig-Kröger A, Serrano-Gómez D, Caparrós E, et al. Regulated expression of the pathogen receptor dendritic cell-specific intercellular adhesion molecule 3 (ICAM-3)-grabbing nonintegrin in THP-1 human leukemic cells, monocytes, and macrophages. J Biol Chem. 2004;279(24):25680–25688. doi:10.1074/jbc.M311516200
33. Azad AK, Torrelles JB, Schlesinger LS. Mutation in the DC-SIGN cytoplasmic triacidic cluster motif markedly attenuates receptor activity for phagocytosis and endocytosis of mannose-containing ligands by human myeloid cells. J Leukoc Biol. 2008;84(6):1594–1603. doi:10.1189/jlb.0308192
34. Barreiro LB, Ben-Ali M, Quach H, et al. Evolutionary dynamics of human Toll-like receptors and their different contributions to host defense. PLoS Genet. 2009;5(7):e1000562. doi:10.1371/journal.pgen.1000562
35. Tailleux L, Pham-Thi N, Bergeron-Lafaurie A, et al. DC-SIGN induction in alveolar macrophages defines privileged target host cells for mycobacteria in patients with tuberculosis. PLoS Med. 2005;2(12):e381. doi:10.1371/journal.pmed.0020381
36. Boily-Larouche G, Zijenah LS, Mbizvo M, et al. DC-SIGN and DC-SIGNR genetic diversity among different ethnic populations: potential implications for pathogen recognition and disease susceptibility. Hum Immunol. 2007;68(6):523–530. doi:10.1016/j.humimm.2007.02.002
37. Vidal SM, Malo D, Vogan K, et al. Natural resistance to infection with intracellular parasites: isolation of a candidate for Bcg. Cell. 1993;73(3):469–485. doi:10.1016/0092-8674(93)90135-D
38. Gruenheid S, Pinner E, Desjardins M, et al. Natural resistance to infection with intracellular pathogens: the nramp1 protein is recruited to the membrane of the phagosome. J Exp Med. 1997;185(4):717–730. doi:10.1084/jem.185.4.717
39. Serrano-Gómez D, Dominguez-Soto A, Ancochea J, et al. Dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin mediates binding and internalization of aspergillus fumigatus conidia by dendritic cells and macrophages. J Immunol. 2004;173(9):5635–5643. doi:10.4049/jimmunol.173.9.5635
40. Quesniaux VJ, Nicolle DM, Torres D, et al. Toll-like receptor 2 (TLR2)-dependent-positive and TLR2-independent-negative regulation of proinflammatory cytokines by mycobacterial lipomannans. J Immunol. 2004;172(7):4425–4434. doi:10.4049/jimmunol.172.7.4425
41. Lu B, Rutledge BJ, Gu L, et al. Abnormalities in monocyte recruitment and cytokine expression in monocyte chemoattractant protein 1-deficient mice. J Exp Med. 1998;187(4):601–608. doi:10.1084/jem.187.4.601
42. Li HT, Zhang TT, Zhou YQ, et al. SLC11A1 (formerly NRAMP1) gene polymorphisms and tuberculosis susceptibility: a meta-analysis. Int J Tuberc Lung Dis. 2006;10(1):3–12.
43. Blackwell JM, Jamieson SE, Burgner D. HLA and infectious diseases. Clin Microbiol Rev. 2009;22(2):370–385. doi:10.1128/CMR.00048-08
44. Ates O, Dalyan L, Müsellim B, et al. NRAMP1 (SLC11A1) gene polymorphisms that correlate with autoimmune versus infectious disease susceptibility in tuberculosis and rheumatoid arthritis. Int J Immunogenet. 2009;36(1):15–19. doi:10.1111/j.1744-313X.2008.00814.x
45. Song W, Li D, Tao L, et al. Solute carrier transporters: the metabolic gatekeepers of immune cells. Acta Pharm Sin B. 2020;10(1):61–78. doi:10.1016/j.apsb.2019.12.006
46. Searle S, Blackwell JM. Evidence for a functional repeat polymorphism in the promoter of the human NRAMP1 gene that correlates with autoimmune versus infectious disease susceptibility. J Med Genet. 1999;36(4):295–299. doi:10.1136/jmg.36.4.295
47. Berrington WR, Hawn TR. Mycobacterium tuberculosis, macrophages, and the innate immune response: does common variation matter? Immunol Rev. 2007;219(1):167–186. doi:10.1111/j.1600-065X.2007.00545.x
48. Selvaraj P, Alagarasu K, Harishankar M, et al. Cytokine gene polymorphisms and cytokine levels in pulmonary tuberculosis. Cytokine. 2008;43(1):26–33. doi:10.1016/j.cyto.2008.04.011
49. Ben-Ali M, Barbouche M-R, Bousnina S, et al. Toll-like receptor 2 Arg677Trp polymorphism is associated with susceptibility to tuberculosis in Tunisian patients. Clin Diagn Lab Immunol. 2004;11(3):625–626. doi:10.1128/CDLI.11.3.625-626.2004
50. Velez DR, Wejse C, Stryjewski ME, et al. Variants in toll-like receptors 2 and 9 influence susceptibility to pulmonary tuberculosis in Caucasians, African-Americans, and West Africans. Hum Genet. 2010;127(1):65–73. doi:10.1007/s00439-009-0741-7
51. Newport MJ, Allen A, Awomoyi AA, et al. The toll-like receptor 4 Asp299Gly variant: no influence on LPS responsiveness or susceptibility to pulmonary tuberculosis in The Gambia. Tuberculosis. 2004;84(6):347–352. doi:10.1016/j.tube.2004.02.001
52. Olesen R, Wejse C, Velez DR, et al. DC-SIGN (CD209), pentraxin 3 and vitamin D receptor gene variants associate with pulmonary tuberculosis risk in West Africans. Genes Immun. 2007;8(6):456–467. doi:10.1038/sj.gene.6364410
53. Khor CC, Chapman SJ, Vannberg FO, et al. A Mal functional variant is associated with protection against invasive pneumococcal disease, bacteremia, malaria and tuberculosis. Nat Genet. 2007;39(4):523–528. doi:10.1038/ng1976
54. Jones BW. Differential roles of Toll-like receptors in the elicitation of proinflammatory responses by macrophages. Ann Rheum Dis. 2001;60(Suppl 3):6–12. doi:10.1136/ard.60.1.6
55. Fitness J, Hill A, Fine P, et al. Large-scale candidate gene study of leprosy susceptibility in the Karonga district of northern Malawi. Am J Trop Med Hyg. 2004;71(3):330–340. doi:10.4269/ajtmh.2004.71.330
56. Chensue SW, Warmington KS, Allenspach EJ, et al. Differential expression and cross-regulatory function of RANTES during mycobacterial (type 1) and schistosomal (type 2) antigen-elicited granulomatous inflammation. J Immunol. 1999;163(1):165–173. doi:10.4049/jimmunol.163.1.165
57. van Crevel R, Ottenhoff TH, Van Der Meer JW. Innate immunity to Mycobacterium tuberculosis. Clin Microbiol Rev. 2002;15(2):294–309. doi:10.1128/CMR.15.2.294-309.2002
58. Saukkonen JJ, Bazydlo B, Thomas M, et al. β-Chemokines Are Induced by Mycobacterium tuberculosis and Inhibit Its Growth. Infect Immun. 2002;70(4):1684–1693. doi:10.1128/IAI.70.4.1684-1693.2002
59. Ben-Selma W, Harizi H, Bougmiza I, et al. Polymorphisms in the RANTES Gene Increase Susceptibility to Active Tuberculosis in Tunisia. DNA Cell Biol. 2011;30(10):789–800. doi:10.1089/dna.2010.1200
60. Lin Y, Gong J, Zhang M, et al. Production of monocyte chemoattractant protein 1 in tuberculosis patients. Infect Immun. 1998;66(5):2319–2322. doi:10.1128/IAI.66.5.2319-2322.1998
61. Flores-Villanueva PO, Ruiz-Morales JA, Song C-H, et al. A functional promoter polymorphism in monocyte chemoattractant protein–1 is associated with increased susceptibility to pulmonary tuberculosis. J Exp Med. 2005;202(12):1649–1658. doi:10.1084/jem.20050126
62. Thye T, Nejentsev S, Intemann CD, et al. MCP-1 promoter variant −362C associated with protection from pulmonary tuberculosis in Ghana, West Africa. Hum Mol Genet. 2009;18(2):381–388. doi:10.1093/hmg/ddn352
Older women and women with disabilities are underrepresented in global data on violence against women. Credit: WHO/Kiana Hayeri
UNITED NATIONS, Mar 29 2024 (IPS) – Older women and women with disabilities experience abuse that is unique to their demographics, yet they are underrepresented in national and global databases, according to findings shared by the World Health Organization (WHO).
On Wednesday, WHO and UN-Women released two new briefs, the first in a series that will discuss neglected forms of violence, including gender-based violence. The two briefs, titled Measuring violence against older women and Measuring violence against women with disability, investigate the types of violence that these groups face through the data available. Through reviewing existing studies into violence against women, the research team was able to synthesize the information available on this topic and its scope across different countries.
As was noted by Dr. Lynnmarie Sardinha, Technical Officer at WHO and the UN Special Programme on Human Reproduction (HRP) for Violence against Women Data and Measurement, and author of the briefs. The limited data on older women and women with disabilities undermines the ability of programmes to meet their needs. “Understanding how diverse women and girls are differently affected, and if and how they are accessing services, is critical to ending violence in all its forms.”
One in three women is affected by gender-based violence in these forms. For older women—aged 60 years and over—and women with disabilities, they are also subjected to other forms of abuse and neglect, usually at the hands of caregivers, family members, or healthcare institutions such as nursing homes. Examples of this include controlling behaviors such as withholding medicine and assistive devices, and financial abuse. Though these forms of neglect and abuse have been observed, the studies that the briefs reviewed seemed to focus more on intimate partner violence through physical and sexual abuse. The briefs acknowledge, however, that violence against women should not only be exemplified by intimate partner violence. The prevalence of this example hints at further nuances that are not sufficiently captured in the studies due to their limitations.
Violence against older women can manifest in other ways as they and their partners/perpetrators age. Although women aged 15–49 are at higher risk of intimate partner and sexual violence, older women are still likely to experience it, and this can shift towards other forms of abuse, such as neglect, economic abuse, and psychological abuse. The brief on older women reveals, however, that there is limited data to definitively state its prevalence. This is particularly the case for low- and middle-income countries; the data that was compiled for this brief comes largely from high-income countries, a gap that the reports are aware of. Older women are represented in only ten percent of the data on violence against women.
Only 6 percent of the studies reviewed for women with disabilities included measures of violence specific to this group. The lack of questions specific to this demographic indicates that they are, perhaps unconsciously, unaccounted for when measuring the scale of violence against women. Data collection procedures may not be designed to accommodate women with disabilities or prevent them from self-reporting, such as deaf or hard-of-hearing women who are unable to participate in surveys conducted through the telephone.
The briefs also suggest that women who live with lifelong disrespect and neglect may not recognize the specific forms of violence, which could account for fewer instances being reported. This could also apply to older women, where surveying and reporting mechanisms are geared towards women of reproductive age, especially in low- and middle-income countries.
This may also speak of socio-cultural attitudes towards violence against older women that are steeped in ageism, harmful stereotypes, and discriminatory cultural norms that prevent them from sharing their experiences.
The WHO briefs make several recommendations to address the evidence gaps. Among them are extending the age limit for survey participation and incorporating questions that relate to different types of violence. Data collection should also account for cultural-specific contexts of violence and abuse across different countries. Women with disabilities should be consulted in research at every stage when designing surveys targeted at them, which will allow for a broader spectrum of disabilities to be accounted for.
Children eating and drinking at the Children’s House in Idlib. Abandoned children is a growing issue in the region. Credit: Sonia Al-Ali/IPS
IDLIB, Syria, Mar 27 2024 (IPS) – Wael Al-Hassan was returning from work in the Syrian city of Harim when he heard the sound of a baby crying.
He was returning from work on December 10, 2023. He stopped momentarily, turned on his mobile phone flashlight to investigate, and spotted a baby girl, around one month old, wrapped in a white blanket, lying by the roadside.
He felt saddened by the infant’s condition and said, “She was crying loudly, and I saw scratches on her face from cat or dog claws. I then carried her in my arms and took her home, where my wife breastfed her, changed her clothes, and took care of her.”
The phenomenon of abandoning newborns is increasing in northern Syria, where individuals leave their newborns in public parks or alongside roads, then leave the area. Passersby later find the infants, some of them dead from hunger or cold.
Al-Hassan said that the next morning, he handed the baby girl over to the police to search for her family and relatives.
Social Rejection
Social worker Abeer Al-Hamoud from the city of Idlib, located in northern Syria, attributes the primary reason for some families abandoning their children to the widespread poverty and high population density in the province. Additionally, there is fear of the security situation (the area is not in the control of the Syrian regime and is often under attack), the prevalence of divorces, and spouses abandoning their families after traveling abroad.
Al-Hamoud also points out another reason, which is the spread of the phenomenon of early marriage and marrying girls to foreign fighters who came from their countries to Syria to participate in combat. Under pressure from their families, wives often have to abandon their children after their husband’s death, sudden disappearance, or return to their homeland, especially when they are unable to care for them or provide for them financially. Moreover, these children have no proper documentation of parentage.
Furthermore, Al-Hamoud mentions another reason, which is some women are raped, leading them to abandon their newborns out of fear of punishment from their families or societal stigma.
Al-Hamoud warns that the number of abandoned children is increasing and says there is an urgent need to find solutions to protect them from exploitation, oppression, and societal discrimination they may face. She emphasizes that the solutions lie in returning displaced persons to their homes, improving living conditions for families, raising awareness among families about the importance of family planning, and launching campaigns to integrate these children into society.
Alternative Families
It’s preferable for members of the community to accept these children into their families, but they face difficulties in registering the births.
Thirty-nine-year-old Samaheer Al-Khalaf from the city of Sarmada in northern Idlib province, Syria, sponsored a newborn found abandoned at a park gate, and she welcomed him into her family.
She says, “After 11 years of marriage to my cousin, we were not blessed with children, so we decided to raise a child found in the city at the beginning of 2022.”
Al-Khalaf observes that the Islamic religion’s prohibition on “adoption” prevents her from registering the child under her name in the civil registry. Additionally, she cannot go to areas controlled by the Syrian regime to register him due to the presence of security barriers.
She says, “I fear for this child’s future because he will remain of unknown lineage. He will live deprived of his civil rights, such as education and healthcare, and he won’t be able to obtain official documents.”
Children’s House Provides Assistance
With the increasing numbers of children of unknown parentage, volunteers have opened a center to receive and care for the children abandoned by their families.
Younes Abu Amin, the director of Children’s House, says, “A child of unknown parentage is one who was found and whose father is unknown, or children whose parentage has not been proven and who have no provider.”
“The organization ‘Children’s House’ opened a center to care for children separated from their families and children of unknown parentage in the city of Sarmada, north of Idlib,” says Abu Amin. “The number of registered children in the center has reached 267, ranging in age from one day to 18 years. Some have been placed with foster families, while others currently reside in the center, receiving all their needs, including shelter, food, education, and healthcare.”
Upon arrival at the center, Abu Amin notes that the center registers each child in its records, transfers them to the shelter department, and makes efforts to locate their original family or relatives and send them to them or to find a foster family to provide them with a decent life.
Abu Amin explains that the center employs 20 staff members who provide children with care, psychological support, and education. They work to create a suitable environment for the children and support them psychologically to help with emotional support.
He emphasizes that the center survives on individual donations to cover its expenses – which are scarce. There is an urgent need for sufficient support, as the children require long-term care, especially newborns.
A young girl Marah (8) and her brother, Kamal (10), lost their father in the war. Their mother remarried, leaving them to live in a small tent with their grandfather, who forces them to beg and sell tissues, often leaving them without food for days.
Consequently, they decided to escape from home. Kamal says, “We used to sleep outdoors, overwhelmed by fear, cold, and hunger, until someone took us to the child center.”
Upon reaching the center, they returned to their studies, played with other children, and each other, just like children with families.
Kamal expresses his wish, “I hope to continue my education with my sister so we can rely on ourselves and escape from a life of injustice and deprivation.”
These children, innocent of any wrongdoing, are often left to fend for themselves, bearing the brunt of war-induced poverty, insecurity, homelessness, instability, and early marriage.
Yamandú Pagliano plans to cross Praia do Cassino, the world’s longest beach, stretching 250 kilometers along Brazil’s southern coast from Uruguay, in his homemade wheeled wind buggy, to highlight the need to address global heating. Credit: Yamandú Pagliano
ROME, Mar 26 2024 (IPS) – Extreme sports are not just for young people. Climate activism isn’t either. Yamandù Pagliano is proof.
The 59-year-old father of two is gearing up for an epic feat. He plans to cross the longest beach on Earth, the Praia do Cassino, stretching from the border of Uruguay 250 kilometres up Brazil’s southern coastline, on his home-made wheeled wind buggy.
It’s a massive challenge both in physical and mental terms and one that brings multiple risks with it, including the danger of getting lost, crashing, or being swept out into the sea if the weather turns nasty.
But the Montevideo native has a special motive for taking it on.
Organizers want to highlight the “institutional indifference” to the climate crisis at all levels of government, promote sustainable transport and tourism, draw attention to the need for more cycle paths, especially in southern Italy, and make a loud appeal for peace around the world
Pagliano is a member of Parents for Future (PFF), a global network of citizens concerned about the climate crisis set up to support and echo the calls made by the young people of the Fridays for Future (FFF) movement.
When he takes on the Praia do Cassino challenge, he’ll be flying the Parents for Future flag on the mast of his wind buggy to highlight the need to address global heating.
“It’s going to be a PFF challenge,” Pagliano told IPS.
“My involvement in PFF started after my daughter joined FFF. Soon I was in Parents for Future Latin America (PFF LATAM) and then I helped to set up PFF Uruguay.
“I hope all the detailed stories of the crossing will help people become aware of the climate crisis, biodiversity loss and pollution. I’ll probably find dead animals and plastic garbage on the beach and face extreme weather events”.
Pagliano knows that the climate crisis is no longer a distant problem for future generations as people in Uruguay have been faced with the consequences first hand, as seen with last year’s severe drought that caused dramatic water shortages.
“In Uruguay the winters are not as cold as they used to be, and summers are a lot hotter,” he said.
“We have had big floods, with houses carried down the coast, and recently we had the biggest drought in our history, with almost no water coming out of the tap”.
Fittingly for an initiative that seeks to show the need for sustainability, Pagliano made his windcar out of reused material, welding together pipes he picked up from a scrap yard, while the sail is second-hand.
“There’ll be no phone signal in the middle section of the beach and I’ll be on my own for quite some time,” said Pagliano, who works in construction.
“I will be completely isolated. You have to be ready for every eventuality.
“Depending on the wind, it could take two or three days.
“It could take just one day with an early departure in good conditions, with the wind blowing in the perfect direction and at the perfect strength.
“It gets tiring physically after a while, but the adrenalin keeps you pumped up.
“It’s a good way to highlight the need to be sustainable.
“It’s a natural sport. There’s no contamination. No carbon footprint.
“I’ll do the crossing first and then go public if I’m successful, like Gagarin,” he quipped.
He is not the only parent harnessing renewable energy to draw attention to the need for climate action.
On the other side of the world, the Italian section of PFF is getting ready for the Running For Future, Cycling For Peace – a bike event which, fittingly for the land of the Giro d’Italia, is split into stages.
The ‘race’ starts in Rome’s Piazza del Popolo on May 10 and features 16 stages over nine days, roughly following the Via Francigena pilgrimage route southwards to end in Lecce on May 19.
Each stage will be used to focus on a specific aspect of the ecological crisis, such as air pollution, urban sprawl and the problems created by intensive livestock farming, while at the same time showing how they are all interconnected.
Among other things, organizers want to highlight the “institutional indifference” to the climate crisis at all levels of government, promote sustainable transport and tourism, draw attention to the need for more cycle paths, especially in southern Italy, and make a loud appeal for peace around the world.
The Italian section of Parents for Future gears up for “Running For Future, Cycling For Peace” — a multi-stage cycling event starting in Rome’s Piazza del Popolo on May 10th and ending in Lecce on May 19th, following the Via Francigena route. Credit: Paul Virgo
“We chose a cycling initiative because the bicycle has become a symbol of ecology,” said Maria Santarossa of Parents for Future Italia. “It is a clean means of transport, which enables you to stay fit and be in direct contact with nature.
“We chose a pilgrims’ path because we can consider it an emblem of the beauty of nature and it’s a way to remind ourselves that we must take care of beauty.
“We have involved many other movements, associations, committees, and networks because we want people to know that many of us have the same objectives regarding the very serious climate and environmental crisis that is present in everyone’s lives”.
It is free to take part in the event, although participants have to cover their own accommodation and food expenses.
It is the second such event. The first took place in 2021, going from Rome northwards along the Via Francigena to Milan for the PreCOP26 conference that was held there.
That was such a success that it inspired the Polish section of Parents for Future to stage a climate grand tour of its own.
Each national PFF group is autonomous and does its own thing, campaigning on the issues that are most appropriate given the local situation.
PFF Italia, for example, is currently engaged in a major campaign to convince consumers to switch to utility companies whose electricity comes only from renewable sources.
There is also an umbrella group, Parents for Future Global (PFFG), which, among other things, is campaigning to support the drive for a Fossil Fuel Non-Proliferation Treaty.
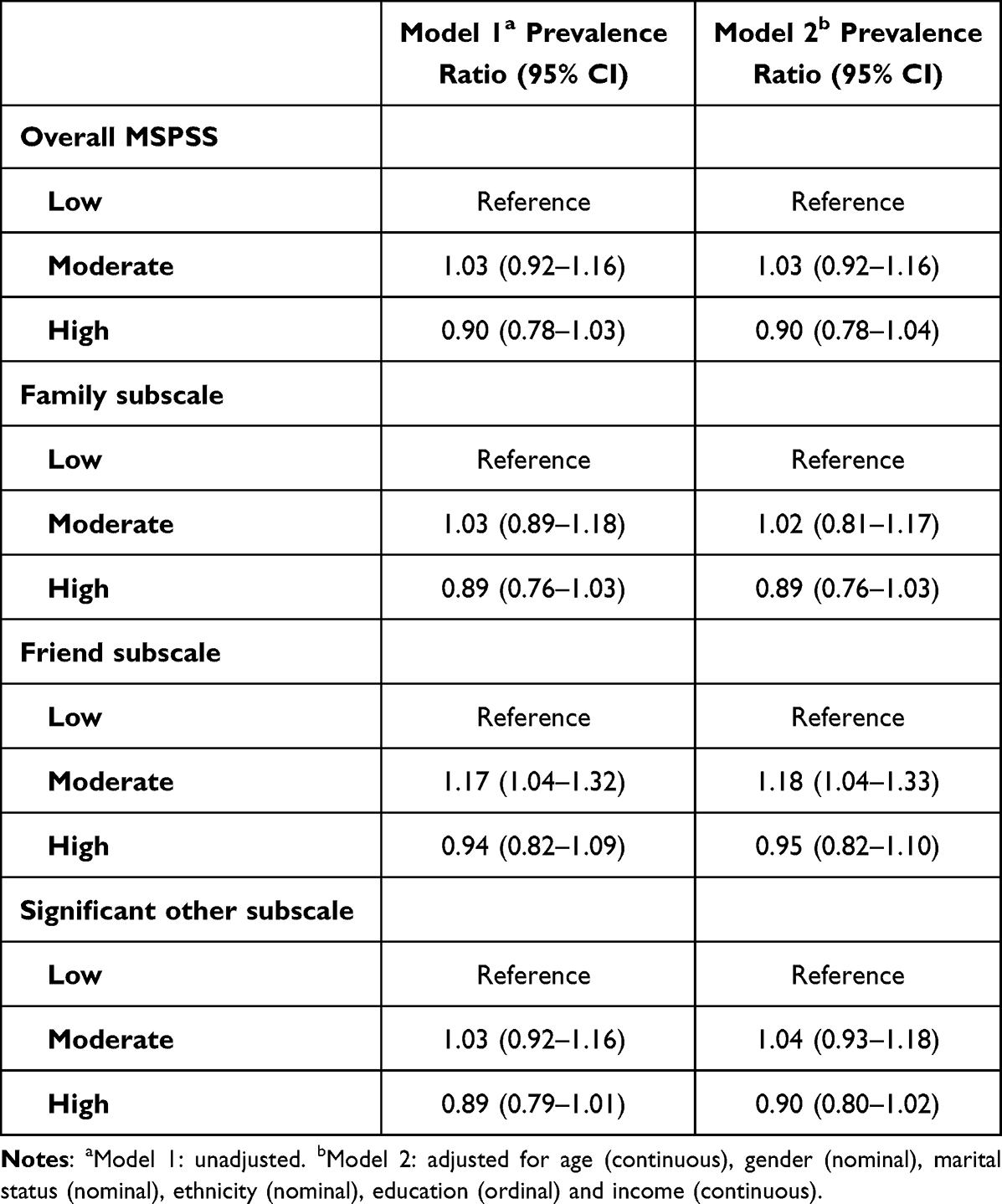
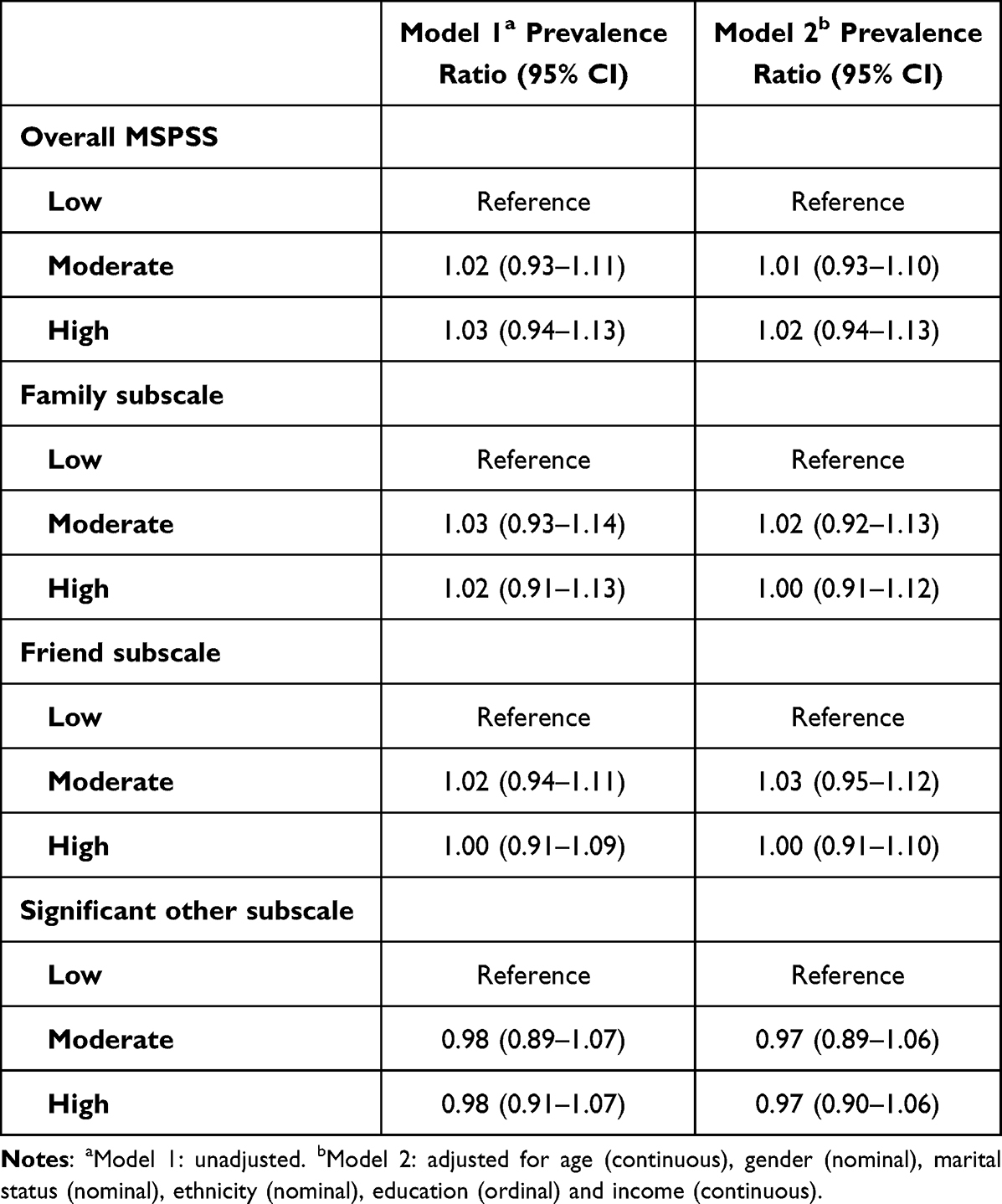
Hypertension is the most common risk factor for cardiovascular diseases (CVDs).1 Globally it affects about 40% of the population and causes approximately 7.6 million deaths every year.2 Despite the availability of effective interventions, including antihypertensive medication only 20% with hypertension have well-controlled blood pressure.3 In Nepal, high blood pressure was the leading cause of CVDs in 20174 where 20–30% of adult women and men have hypertension.5 The high proportion of untreated (89% of those aware) and uncontrolled (96% of those on treatment) hypertension in Nepal,5 jeopardizes the government’s commitment to reduce CVDs.6
Management of hypertension may require substantial efforts, including adherence to antihypertensive medications, monitoring of blood pressure, frequent follow-up with healthcare providers, weight reduction, physical activity, healthy diet, and avoidance of alcohol and tobacco use.7 Patients with chronic conditions like hypertension often need social and family support to optimally manage their ailment.8,9 Social support is a multidimensional concept often defined as activities and relationships that individuals receive and provide to each other within their social networks.10 A large and diverse social support network can boost self-esteem and provide better access to information and resources.11,12 Health-related interactions with friends and family promote healthy behaviors including adherence to treatment.13–16 Feeling connected with others and the awareness that support is available when needed are important for positive health outcomes.17–19 Positive social relationships help patients cope with illness associated stress promoting better prognosis.20 Social support may protect patients from complications by helping with health management,16 or by encouraging healthy behaviors.21 However, the evidence on the role of social support in the management of chronic conditions is inconclusive. Studies have shown a positive,22–26 null27,28 and an inverse association29 between social support and management of chronic conditions. The role of social support in the management of chronic conditions like hypertension has not been sufficiently explored in the Nepalese context. One study from Nepal reported a positive association between social support and self-care for hypertension.30 In this study, we explored how perceived social support influences medication adherence and control of high blood pressure. The results from this study could serve to inform targeted community-based interventions according to the level of social support received by hypertension patients for controlling hypertension.
Materials and Methods
Study Setting
The study was conducted in Budhanilkantha municipality, Kathmandu, Nepal. The municipality has nearly 150,000 inhabitants31 and has 11 public health facilities that provide primary health care and tertiary care is provided by the private and public hospitals nearby. Hypertension prevalence of the urban areas such as Budanilakantha (25.2%) is similar to the national average (24.5%).5
Study Design and Population
Cross-sectional data collected at baseline from 1252 hypertensive individuals enrolled in a cluster randomized trial (Registration no: NCT05292469). The detailed trial methods are published elsewhere.32 Trained enumerators identified participants seeking support from health workers and volunteers, screened for eligibility, and obtained written informed consent after explaining the trial objectives. Eighteen years and older individuals with established hypertension diagnosis (systolic BP ≥140 mmHg and/or diastolic BP ≥ 90 mmHg on at least two consecutive visits or using antihypertensive medication) and able to respond to the questions were recruited. Pregnant women were excluded.
Data Collection
Trained enumerators collected baseline data from 2 May 2022 through 7 November 2022 using an android operating system tablet installed with KOBO toolbox electronic data collection platform. The questionnaire was pretested. Participants were shown picture cards with examples of physical activity and commonly used utensils for drinking alcohol to ensure accurate measurements.5
Outcomes
Hypertension control
An “Omron” digital instrument was used to measure blood pressure three times in a resting position and the mean of the last two measurements was registered. Participants with systolic and diastolic blood pressure less than 140 and 90 mmHg were categorized as controlled hypertension others as uncontrolled hypertension.
Medication adherence
We administered an eight-item Morisky Medication Adherence Scale (MMAS-8) (Supplementary Table 1).33 The scale is widely used to measure medication adherence and is reported to have good reliability (α = 0.83) and validity with sensitivity of 93% and specificity of 53% for low adherence.34 The first seven questions have responses YES coded as 0 and NO coded as 1. The code of the fifth question was reversed and the responses to the eighth question were re-coded from 2 to 0.75, 3 to 0.5, 4 to 0.25, 5 to 0 during the analysis. Responses to all eight questions were added to get MMAS, and a score above 6 was considered good adherence and ≤6 poor adherence.35
Exposures
Modified multidimensional scale of perceived social support (MSPSS) was used to measure the adequacy of support participants received from family, friends, and significant others (Supplementary Table 2).36 Twelve questions with answers on a 5-point Likert scale (1 = strongly disagree; 5=strongly agree) were asked to the participants. Cronbach’s alpha has been estimated to be 0.92, 0.85, 0.85 and 0.86 for total and family, friends, and significant others subscales, respectively.36 The MSPSS was translated into Nepali by SB and was reviewed by the study clinician. During training, the wording of the questions was discussed with the enumerators from the study community. The enumerators pretested the questions, and any problems encountered in administering the questions were discussed and addressed before finalizing the tool. The internal consistency reliability for the MSPSS was found good with a Cronbach’s alpha of 0.91 for the overall score and 0.87, 0.91, and 0.84 for family, friend, and significant other subscales, respectively. The overall MSPSS ranged between 12 and 60, high scores indicate high perceived social support. The subscale scores were calculated by adding the responses for related questions. During analysis, scores were divided by the number of questions included in each scale resulting in scores ranging between 1 and 5. The scores were categorized as low (1.00–3.59), moderate (3.60–4.59), and high (4.60–5.00) support.
Covariates
The covariates included in the model were age (continuous), gender (female/male), ethnicity (Brahmin Chettri/Newars/Tamang, Sherpa, Rai, Gurung, Magar/Dalits), current marital status (unmarried/married), education (illiterate/primary/secondary/high school and above), and per capita annual income in US dollars (continuous). Other variables considered were occupation status (unemployed/paid employment), self-reported diabetes status (status not known/non-diabetic/diabetic), years since hypertension diagnosis, and prescribed antihypertensives (Yes/No). The diet quality questionnaire assessed 32 different food groups participants ate in the 24 hours preceding the survey.37 The global dietary recommendation score (ranging from 0 to 18, higher score means better diet quality) was calculated by subtracting foods recommended to limit from foods recommended as healthy and adding nine. It measures adherence to a healthy diet protective against non-communicable diseases.37 Fruits and vegetable scores ranged from 0 to 6, with scores of <3 indicate the likelihood of eating less than 400 grams of fruits and vegetables.37 Daily salt intake was asked and categorized as ≤10 grams, 11–15 grams, and >15 grams. A global physical activity questionnaire38 was used to calculate metabolic equivalents of task (METs) minutes per week categorizing <600 METs (inadequate) and ≥600 (adequate). Body mass index of <25 kg/m2 and ≥25 kg/m2 was categorized as normal weight and overweight, respectively.39 Standard drinks per week were calculated by asking current alcohol drinkers about drinking frequency and amount of different types of alcohol5 and categorizing into non-drinkers (≤1 standard drink per month), moderate drinkers (<3 standard drinks per week), and high drinkers (≥3 standard drinks per week). Tobacco use including both smoked and chewed was categorized as never, and ever users.
Statistical Analysis
Descriptive statistics were presented as frequencies and percentages for categorical variables and mean and standard deviation for continuous variables. Confounders included in the model were identified a priori by constructing a directed acyclic graph (DAG)40 to assess the association between MSPSS and controlled hypertension and adherence to antihypertensives. We fitted Poisson regression to assess the association between MSPSS and hypertension control. Prevalence ratios and corresponding 95% confidence intervals were estimated. Two models were fitted, unadjusted (model 1) and adjusted for age, gender, ethnicity, marital status, education, and income (model 2). Similar models were also fitted to assess the association between MSPSS and adherence to antihypertensives. Based on the DAGs, mediators such as dietary, lifestyle and clinical factors were not included in the model. All analyses were performed with Stata 18.41
Ethics
We have obtained ethical approval from Nepal Health Research Council (Protocol number: 682/2021) approved on 24 December 2021 and Regional Committee for Medical and Health Research Ethics, Norway (Reference number: 399479) approved on 22 February 2022. We have adhered to the declaration of Helsinki throughout the research process.
Results
The mean age of study participants was 57.5 years, and 60% were females. Table 1 shows the distribution of sociodemographic, lifestyle and clinical factors by categories of overall MSPSS. Participants with high social support were generally younger and more often males, married, belonged to Brahmin/Chettri ethnicity, were highly educated, had paid employment, had high per capita income, physically active, overweight, drink <3 standard drinks per week, current tobacco users, consumed daily <10 grams salt, had high global dietary requirement score, consumed less fruits and vegetables, were diabetic, were prescribed more than three antihypertensives, had shorter duration since diagnosis of hypertension and had poorly controlled blood pressure.
Table 1 Socioeconomic, Lifestyle, and Clinical Factors by Levels of Overall MSPSS
Out of the 1252 participants 914 (73%) reported receiving moderate to high overall MSPSS. Figure 1 shows that the high proportion of individuals with controlled hypertension and good adherence to antihypertensives were in moderate MSPSS category (58.7%) and high MSPSS category (74.3%), respectively. The distribution of MSPSS subscales by status of hypertension control and adherence to antihypertensives are shown in Supplementary Tables 3 and 4.
Figure 1 Proportion controlling hypertension and good adherence to antihypertensives in categories of overall MSPSS.
Overall, there was no significant association between social support and controlled hypertension (Table 2). However, individuals who received a moderate level of social support from friends exhibited 1.18 (95% CI, 1.04–1.33) times higher prevalence of controlled hypertension compared to those receiving low support.
Table 2 Association Between MSPSS and Controlled Hypertension
Further, Table 3 shows that there was no association between social support and prevalence of adherence to antihypertensives, and unadjusted and adjusted estimates were very similar.
Table 3 Association Between MSPSS and Adherence to Antihypertensives
Discussion
In this study, we explored how perceived social support received by hypertension patients affected adherence to antihypertensives and control of hypertension. Our analysis showed that 73% of participants received moderate to high social support. Despite anticipating a positive association between MSPSS and controlled hypertension, we found that participants in the high MSPSS category had the lowest proportion of hypertension control (51%) while the proportion of good adherence did not differ between MSPSS categories. There was no association between MSPSS and adherence to antihypertensives and controlled hypertension except for friend’s sub-scale where higher prevalence of controlled hypertension was observed in moderate social support compared to low.
Very few studies from Nepal have reported on the relationship between social support and hypertension control and adherence to antihypertensives. Our finding that hypertensive patients receive high social support is in line with previous studies from Ethiopia42 and Turkey,43 but not with studies from Nepal30 and Malaysia.44 Previous studies from high-income countries agree with our findings that social support is neither associated with hypertension control45,46 (Spain/Korea) nor with adherence to antihypertensives28 (USA). However, a positive association between social support with controlled hypertension has been reported by a study from Vietnam47 and with adherence to antihypertensives by studies from India,23 China48 and Turkey.43 The inconsistent findings could be due to discrepancies in how social support was measured. A few studies have used MSPSS30,43,49 to measure social support but their outcome measure was self-efficacy to hypertension rather than hypertension control while those with hypertension control as the outcome measured social support differently by measuring frequency of visits by friends and family,45 social support inventory,28 self-esteem and belongingness23 and availability of informational, emotional, practical support.27,46,48
The quality and type of social support received has a bearing on adherence to medication and hypertension control.13 Practical support received for household chores lowered the risk of uncontrolled hypertension in Vietnam,47 whereas financial support from friends was harmful to adherence to medication in China.48 Therefore, nuanced studies disentangling the mechanisms through which social support the process of hypertension management are needed.
The reason hypertensive patients received high social support in our study may be due to availability of family members rendered by large family size in our sample. Despite the high level of support received by participants, we did not observe a significant positive association between MSPSS and hypertension control and adherence to antihypertensives. This may be due to a tendency of parents not wanting to burden children with their health concerns.50 Also, nearly 33% of the participants were illiterate and their friends and family are likely similar. Even when support is available, a poorly educated social network may not be capable of imparting the informational support needed to adhere to treatment and thereby control hypertension.51
Another explanation to the discrepancy of our results with other studies might be differences in individual and cultural characteristics. A large proportion of unemployed individuals may have skewed our sample towards a higher level of social support. People value financial support and therefore financially dependent individuals may report receiving high support. Also, the threshold for expected support may vary between different societies, for example in a patriarchal society like Nepal, women as primary care takers in the family may have a lower threshold of social support, whereas elderly men who command more respect in society may have a higher threshold.
We found a numerically higher prevalence of controlled hypertension among those receiving moderate support from friends but not those receiving higher support. A possible explanation might be that those receiving higher support spend longer time with friends and are thereby more exposed to peer pressure for unhealthy behaviour which was indicated as a barrier for effective control of blood pressure in our formative study in the same population.52 Reverse causation can also not be ruled out, as those with poorly controlled blood pressure may be receiving higher support from friends.
Strengths and Limitations
Our study adds to the sparse literature examining this association using validated instruments such as MSPSS and MMAS-8. We recruited hypertension patients from the community (not hospital) ensuring a representative sample. Since the outcome of interest (controlled hypertension and good adherence to antihypertensives) was common (larger than 10%), we fitted a modified Poisson regression model which gives a better approximation of the risk than overestimated odds ratio.53 This study is not without limitations. The cross-sectional design may have masked the true association between social support and controlled hypertension, as it is possible that individuals with uncontrolled hypertension or with poor adherence were receiving more support. Given the numerous statistical tests we conducted, it is essential to interpret any significant findings with caution to mitigate the risk of drawing false-positive conclusions. The findings are based on data from one urban municipality, which might limit the generalizability of our findings to the general hypertensive population in Nepal. Social support as a construct would be better explored using a mixed-method approach as the MSPSS tool does not capture the type of social support received.49 The reliance on self-reported data on medication and social support may have resulted in recall bias skewing the data in the direction of social desirability leading to over-estimated adherence and social support.
Conclusion
This study shows that hypertensive patients in Nepal in general have good social support.
However, social support was not foremost for medication adherence and control of hypertension. Future interventions for example digital technologies that facilitate remote monitoring and communication with care providers54 should nurture the high level of social support received by the hypertension patients. However, contextual studies to delineate the mechanisms through which social support can augment adherence to treatment, healthy behaviour, routine monitoring, and follow-up for hypertension management are needed for social support to have precedence in future interventions for hypertension patients.
Abbreviations
CVDs, Cardiovascular diseases; DAG, Directed acyclic graph; METs, Metabolic equivalents of task; MMAS, Morisky medication adherence scale; MSPSS, Multidimensional scale of perceived social support.
Data Sharing Statement
Data cannot be shared publicly for ethical reasons but are available on reasonable request to Ms Sanju Bhattarai and Dr Abhijit Sen.
Acknowledgments
We would like to thank all the participants in the study. We would like to thank field researchers Ms Apsara Basnet, Ms Geeta Tripathi, Ms Pema Tamang, Ms Prasoon Pandey, Mr Kishore Adhikari, Ms Sarita Bhandari, Ms Supriya Kharel.
The MMAS-8 Scale, content, name, and trademarks are protected by US copyright and trademark laws. Permission for use of the scale and its coding is required. A license agreement is available from MMAR, LLC., www.moriskyscale.com.
Funding
This work was supported by Norwegian University of Science and Technology, Trondheim, Norway (Project number 981023100).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Roth GA, Johnson C, Abajobir A, et al. Global, regional, and national burden of cardiovascular diseases for 10 causes, 1990 to 2015. J Am Col Cardiol. 2017;70(1):1–25. doi:10.1016/j.jacc.2017.04.052
2. Lawes CM, Hoorn SV, Rodgers A. Global burden of blood-pressure-related disease, 2001. Lancet. 2008;371(9623):1513–1518. doi:10.1016/S0140-6736(08)60655-8
3. Chow CK. Prevalence, awareness, treatment, and control of hypertension in rural and urban communities in high-, middle-, and low-income countries. JAMA. 2013;310(9):959. doi:10.1001/jama.2013.184182
4. Bhattarai S, Aryal A, Pyakurel M, et al. Cardiovascular disease trends in Nepal – an analysis of global burden of disease data 2017. IJC Heart Vasculature. 2020;30(100602):100602. doi:10.1016/j.ijcha.2020.100602
5. Dhimal MBB, Bhattarai S. Report of non communicable disease risk factors: STEPS survey Nepal 2019. Kathmandu; 2020.
6. Collaborators NC, Stevens GA, Mathers CD. NCD countdown 2030: worldwide trends in non-communicable disease mortality and progress towards sustainable development goal target 3.4. Lancet. 2018;392(10152):1072–1088. doi:10.1016/S0140-6736(18)31992-5
7. Prevention, detection, evaluation, and treatment of high blood pressure the seventh report of the joint national committee on complete report. U.S. Department of Health and Human Services, National Institutes of Health, National Heart, Lung, and Blood Institute, National High Blood Pressure Education Program; Contract No.: 04-5230; 2004.
8. Barth J, Schneider S, von Känel R. Lack of social support in the etiology and the prognosis of coronary heart disease: a systematic review and meta-analysis. Psychosomatic Med. 2010;72(3):229–238. doi:10.1097/PSY.0b013e3181d01611
9. Eom C-S, Shin DW, Kim SY, et al. Impact of perceived social support on the mental health and health-related quality of life in cancer patients: results from a nationwide, multicenter survey in South Korea. Psycho-Oncology. 2013;22(6):1283–1290. doi:10.1002/pon.3133
10. Tardy CH. Social support measurement. Am J Community Psychol. 1985;13(2):187–202. doi:10.1007/BF00905728
11. Scanfeld D, Scanfeld V, Larson EL. Dissemination of health information through social networks: twitter and antibiotics. Am J Infect Control. 2010;38(3):182–188. doi:10.1016/j.ajic.2009.11.004
12. Perry BL, Pescosolido BA. Functional specificity in discussion networks: the influence of general and problem-specific networks on health outcomes. Social Networks. 2010;32(4):345–357. doi:10.1016/j.socnet.2010.06.005
13. DiMatteo MR. Social support and patient adherence to medical treatment: a meta-analysis. Health Psychol. 2004;23(2):207–218. doi:10.1037/0278-6133.23.2.207
14. Trivedi RB, Ayotte B, Edelman D, Bosworth HB. The association of emotional well-being and marital status with treatment adherence among patients with hypertension. J Behavl Med. 2008;31(6):489–497. doi:10.1007/s10865-008-9173-4
15. Cornwell EY, Waite LJ. Social network resources and management of hypertension. J Health Social Behav. 2012;53(2):215–231. doi:10.1177/0022146512446832
16. Gallant MP, Spitze GD, Prohaska TR. Help or hindrance? How family and friends influence chronic illness self-management among older adults. Research on Aging. 2007;29(5):375–409. doi:10.1177/0164027507303169
17. Callaghan P, Morrissey J. Social support and health: a review. J Adv Nurs. 1993;18(2):203–210. doi:10.1046/j.1365-2648.1993.18020203.x
18. Bolger N, Zuckerman A, Kessler RC. Invisible support and adjustment to stress. J Pers Soc Psychol. 2000;79(6):953–961. doi:10.1037/0022-3514.79.6.953
19. Norris FH, Kaniasty K. Received and perceived social support in times of stress: a test of the social support deterioration deterrence model. J Pers Soc Psychol. 1996;71(3):498–511. doi:10.1037/0022-3514.71.3.498
20. Idler EL, Boulifard DA, Contrada RJ. Mending broken hearts. J Health Social Behav. 2012;53(1):33–49. doi:10.1177/0022146511432342
21. Ford CD, Sims M, Higginbotham JC, et al. Psychosocial factors are associated with blood pressure progression among African Americans in the Jackson Heart Study. Am J Hypertens. 2016;29(8):913–924. doi:10.1093/ajh/hpw013
22. Taher M, Abredari H, Karimy M, Abedi A, Shamsizadeh M. The relation between social support and adherence to the treatment of hypertension. J Educ Community Health. 2014;1(3):63–69. doi:10.20286/jech-010348
23. Padhy M, Lalnuntluangi R, Chelli K, Padiri RA. Social support and adherence among hypertensive patients. AJHM. 2016;1(1):33–40.
24. Pan J, Hu B, Wu L, Li Y. The effect of social support on treatment adherence in hypertension in China. Patient Preference Adherence. 2021;15:1953–1961. doi:10.2147/PPA.S325793
25. Osamor PE. Social support and management of hypertension in south-west Nigeria: cardiovascular topic. Cardiovasc J Afr. 2015;26(1):29–33. doi:10.5830/CVJA-2014-066
26. Sabaté E, World Health O. Adherence to long-term therapies: evidence for action. World Health Organization. 2003;194.
27. Adisa R, Olajide OO, Fakeye TO. Social support, treatment adherence and outcome among hypertensive and Type 2 diabetes patients in ambulatory care settings in southwestern Nigeria. Ghana Med J. 2017;51(2):64–77.
28. Hm ST, Quyyumi A, Reilly C, Pemu P, Dunbar S, Dunbar S. The relationship among health beliefs, depressive symptoms, medication adherence, and social support in African Americans with hypertension. J Cardiovasc Nurs. 2019;34(1):44–51. doi:10.1097/JCN.0000000000000519
29. Uchino BN. Understanding the links between social support and physical health: a life-span perspective with emphasis on the separability of perceived and received support. Perspectives Psychol Sci. 2009;4(3):236–255. doi:10.1111/j.1745-6924.2009.01122.x
30. Acharya R, Chaudhary A, Pandey J, Pandey C. Self-Care Management And Its Associated Factors Among Patient With hypertension in Nepal. J Chitwan Med Coll. 2022;12(2):91–96. doi:10.54530/jcmc.637
31. Karki M Budhanilakantha municipality profile; 2019.
32. Bhattarai S, Shrestha A, Skovlund E, Åsvold BO, Mjølstad BP, Sen A. Cluster randomised trial to evaluate comprehensive approach to hypertension management in Nepal: a study protocol. BMJ open. 2023;13(5):e069898. doi:10.1136/bmjopen-2022-069898
33. Morisky DE AA, Krousel-Wood M, Ward HJ, Ward HJ. Predictive validity of a medication adherence measure in an outpatient setting. J Clin Hypertens. 2008;10(5):348–354. doi:10.1111/j.1751-7176.2008.07572.x
34. De M Concurrent and predictive validity of a self-reported measure of medication adherence; 1986.
35. Lee GKY, Wang HHX, Liu KQL, Cheung L, Morisky DE, Wong MCS. Determinants of medication adherence to antihypertensive medications among a Chinese population using morisky medication adherence scale. PLoS One. 2013;8(4):e62775. doi:10.1371/journal.pone.0062775
36. Stewart RC, Umar E, Tomenson B, Creed F. Validation of the multi-dimensional scale of perceived social support (MSPSS) and the relationship between social support, intimate partner violence and antenatal depression in Malawi. BMC Psychiatry. 2014;14(1). doi:10.1186/1471-244X-14-180
38. Global physical activity questionnaire Global Physical Activity Questionnaire (GPAQ). [Internet]. Available from: www.who.int/chp/steps. Accessed March 21, 2024.
39. World Health Organization. GLobal status report on non communicable diseases 2014; 2014.
40. Textor J. Adjustment criteria in causal diagrams: an algorithmic perspective. In Proceedings of the 27th Conference on Uncertainty in Artificial Intelligence (UAI 2011) [press release]; 2011.
41. StataCorp. Stata Statistical Software: Release 18. College Station, TX: StataCorp LLC; 2023.
42. Ademe SAF, Gela D, Gela D. Hypertension self-care practice and associated factors among patients in public health facilities of Dessie town, Ethiopia. BMC Health Serv Res. 2019;19(1):1–10. doi:10.1186/s12913-019-3880-0
43. Rabia Hacihasanoglu Asilar AY, Saglam R, Demirturk Selcuk E, Erduran Y, Sarihan O. The effect of loneliness and perceived social support on medication adherence self-efficacy in hypertensive patients: an example of Turkey. Eur J Cardiovasc Nurs. 2020;38(4):183–190.
44. Nur Syafaah Nazeri NM, Redzuan M, Abdul Mulud Z. Social support and self-care practice among patients with hypertension in a teaching hospital. Malaysian J Med Health Sci. 2021;18(8):281–290.
45. Redondo-Sendino PG-C Á, José RB, Rodríguez-Artalejo F, Rodríguez-Artalejo F. Relationship between social network and hypertension in older people in Spain. Rev Esp Cardiol. 2005;58(11):1294–1301.
46. Jiwon Baek NWH, Chang Kim H, Youm Y, Youm Y. Sex-specific effects of social networks on the prevalence, awareness, and control of hypertension among older Korean adults. J Geriatric Cardiol. 2016;13(7):580–586. doi:10.11909/j.issn.1671-5411.2016.07.005
47. Luu Quang Thuy NHT, Hong Trung L, Huy Tan P, et al. Blood pressure control and associations with social support among hypertensive outpatients in a developing country. Biomed Res Int. 2021;2021:1–10.
48. Bingjie Shen TG, Xinyu D, Chenyang P, Zhao J, Liu Y, Liu Y. Medication adherence and perceived social support of hypertensive patients in China: a community-based survey study. Patient Preference Adherence. 2022;16:1257–1268. doi:10.2147/PPA.S363148
49. Hatah ELK, Ali AM, Mohamed Shah N, Islahudin F, Islahudin F. The influence of cultural and religious orientations on social support and its potential impact on medication adherence. Patient Prefer Adherence. 2015;9:589–596. doi:10.2147/PPA.S79477
50. Acharya TDG, Traille K, Dhakal H. Senior citizens in Nepal: policy gaps and recommendations. Gerontol Geriatr Med. 2023;9:23337214231179902.
51. Isa MD, Shahar SHF, He FJ, Majid HA. Associations of health literacy with blood pressure and dietary salt intake among adults: a systematic review. Nutrients. 2021;13(12):4534.
52. Bhattarai S, Bajracharya S, Shrestha A, et al. Facilitators and barriers to hypertension management in urban Nepal: findings from a qualitative study. Open Heart. 2023;10(2):e002394. doi:10.1136/openhrt-2023-002394
53. Spiegelman D. Easy SAS calculations for risk or prevalence ratios and differences. Am J Epidemiol. 2005;162(3):199–200.
54. Kario K, Nomura A, Harada N, et al. Efficacy of a digital therapeutics system in the management of essential hypertension: the HERB-DH1 pivotal trial. Eur Heart J. 2021;42(40):4111–4122. doi:10.1093/eurheartj/ehab559